Ein Aussterben (auch bekannt als Massenaussterben oder biotische Krise ) ist ein weit verbreiteter und schneller Rückgang der Artenvielfalt auf der Erde . Ein solches Ereignis ist durch einen starken Rückgang der Vielfalt und Häufigkeit mehrzelliger Organismen gekennzeichnet . Es tritt auf, wenn die Aussterberate im Verhältnis zur Hintergrundaussterberate [1] und der Artbildungsrate zunimmt . Schätzungen zur Zahl der großen Massenaussterben in den letzten 540 Millionen Jahren reichen von nur fünf bis über zwanzig. Diese Unterschiede resultieren aus Uneinigkeit darüber, was ein „großes“ Aussterbeereignis darstellt und welche Daten zur Messung der vergangenen Vielfalt ausgewählt wurden.

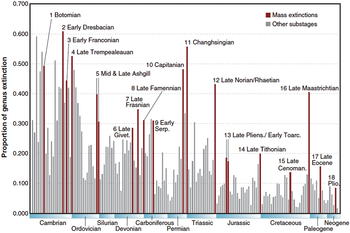
Intensität des Meeressterbens während des Phanerozoikums
%
Vor Millionen von Jahren
Die blaue Grafik zeigt den scheinbaren
Prozentsatz (nicht die absolute Zahl) der Meerestiergattungen, die
in einem bestimmten Zeitraum ausgestorben sind. Es handelt sich nicht um alle Meeresarten, sondern nur um diejenigen, die leicht versteinerbar sind. Die Bezeichnungen der traditionellen „Big Five“-Aussterbeereignisse und des neueren bekannt gewordenen
Massenaussterbens auf den Capitanien sind anklickbare Links. Die beiden Aussterbeereignisse im Kambrium (ganz links) sind prozentual sehr groß, aber in absoluten Zahlen der bekannten Taxa gering, da zu dieser Zeit relativ wenig fossiles Leben produziert wurde.

Inhalt
Die „Big Five“-Massensterbenbearbeiten
In einer bahnbrechenden Arbeit aus dem Jahr 1982 identifizierten Jack Sepkoski und David M. Raup fünf besondere geologische Abschnitte mit übermäßigem Diversitätsverlust. [2] Sie wurden ursprünglich als Ausreißer eines allgemeinen Trends sinkender Aussterberaten während des Phanerozoikums identifiziert, [3] aber als strengere statistische Tests auf die gesammelten Daten angewendet wurden, wurde festgestellt, dass im aktuellen Phanerozoikum Das vielzellige Tierleben hat mindestens fünf große und viele kleinere Massenaussterben erlebt. [4] Die „Big Five“ können nicht so klar definiert werden, sondern scheinen vielmehr die größten (oder einige der größten) eines relativ gleichmäßigen Kontinuums von Aussterbeereignissen darzustellen. [3] Es wird spekuliert, dass es sich um ein früheres (erstes?) Ereignis am Ende des Ediacaran handelte, und allen geht das vermutlich weitaus umfangreichere Massensterben mikrobiellen Lebens während der Sauerstoffkatastrophe zu Beginn des Proterozoikums voraus . [5]
- Ordovizium-Silur-Aussterbeereignisse (Endordovizium oder O–S): 445–444 Ma , kurz vor und am Ordovizium – Silur- Übergang. Es ereigneten sich zwei Ereignisse, die 27 % aller Familien , 57 % aller Gattungen und 85 % aller Arten töteten. [6] Zusammengenommen werden sie von vielen Wissenschaftlern als das zweitgrößte der fünf großen Artensterben in der Erdgeschichte eingestuft, gemessen am Prozentsatz derausgestorbenen Gattungen . Im Mai 2020 deuteten Studien darauf hin, dass die Ursachen des Massensterbens die globale Erwärmung im Zusammenhang mit Vulkanismus und Anoxie seien und nicht, wie zuvor angenommen, Abkühlung und Vereisung . [7] [8] Dies steht jedoch im Widerspruch zu zahlreichen früheren Studien, die die globale Abkühlung als Hauptursache angegeben haben. [9] In jüngster Zeit wurde vermutet, dass die Ablagerung von Vulkanasche der Auslöser für die Verringerung des atmosphärischen Kohlendioxids ist, was zu der in den geologischen Aufzeichnungen beobachteten Vereisung und Anoxie führt. [10]
- Aussterben im Spätdevon : 372–359 Ma , besetzt einen Großteil des Oberdevons bis zum Übergang Devon – Karbon . Das Oberdevon war ein Zeitraum mit hohem Diversitätsverlust, der sich auf zwei Aussterbeereignisse konzentrierte. Das größte Aussterben war das Kellwasser-Ereignis ( Frasnium – Famennium oder FF, 372 Ma), ein Aussterben am Ende des Frasniums, etwa in der Mitte des späten Devon. Dieses Aussterben vernichtete Korallenriffe und zahlreiche tropische benthische (auf dem Meeresboden lebende) Tiere wie kieferlose Fische, Brachiopoden und Trilobiten . Ein weiteres großes Aussterben war das Hangenberg-Ereignis (Devon-Karbon oder DC, 359 Ma), das das Ende des gesamten Devon bedeutete. Dieses Aussterben vernichtete die gepanzerten Placodermfische und führte beinahe zum Aussterben der neu entwickelten Ammonoide . Durch diese beiden kurz aufeinanderfolgenden Aussterbeereignisse wurden insgesamt etwa 19 % aller Familien, 50 % aller Gattungen [6] und mindestens 70 % aller Arten ausgelöscht. [11] Sepkoski und Raup (1982) [2] betrachteten das Aussterbenintervall im späten Devon ( Givetian- , Frasnium- und Famennium-Stadium) zunächst nicht als statistisch signifikant. [2] Unabhängig davon haben spätere Studien die starken ökologischen Auswirkungen der Kellwasser- und Hangenberg-Ereignisse bestätigt. [12]
- Trilobiten waren äußerst erfolgreiche Meerestiere, bis das Aussterben im Perm und der Trias sie alle auslöschte.Perm-Trias-Aussterbeereignis (Endperm): 252 Ma , am Perm – Trias- Übergang. [13] Das größte Aussterben des Phanerozoikums tötete 53 % der Meeresfamilien, 84 % der Meeresgattungen, etwa 81 % aller Meeresarten [14] und schätzungsweise 70 % der Landwirbeltierarten. [15] Dies ist auch das größte bekannte Aussterben von Insekten . [16] Der äußerst erfolgreiche Meeresgliederfüßer, der Trilobit , starb aus. Die Beweise für Pflanzen sind weniger eindeutig, aber nach dem Aussterben wurden neue Taxa dominant. [17] Das „Große Sterben“ hatte eine enorme evolutionäre Bedeutung: An Land beendete es die Vorherrschaft der frühen Synapsiden . Die Erholung der Wirbeltiere dauerte 30 Millionen Jahre, [18] aber die leeren Nischen boten den Archosauriern die Möglichkeit, auf dem Vormarsch zu. In den Meeren sank der Anteil der Tiere, die sich nicht fortbewegen konnten, von 67 % auf 50 %. Das gesamte späte Perm war eine schwierige Zeit, zumindest für das Meeresleben, sogar vor dem Aussterben der P-T-Grenze. Neuere Forschungen haben gezeigt, dass das End-Capitanian-Aussterben , das dem „Großen Sterben“ vorausging, wahrscheinlich ein vom P-T-Aussterben getrenntes Ereignis darstellt; Wenn ja, wäre es größer als einige der „Big Five“-Aussterbeereignisse und verdient möglicherweise einen separaten Platz in dieser Liste unmittelbar vor diesem.
- Trias-Jura-Aussterbeereignis (Endtrias): 201,3 Ma , am Übergang Trias – Jura . Etwa 23 % aller Familien, 48 % aller Gattungen (20 % der Meeresfamilien und 55 % der Meeresgattungen) und 70 bis 75 % aller Arten starben aus. [6] Die meisten nicht-dinosauriischen Archosaurier , die meisten Therapsiden und die meisten großen Amphibien wurden eliminiert, so dass die Dinosaurier nur noch wenig Konkurrenz auf der Erde hatten. Nicht-Dinosaurier-Archosaurier dominierten weiterhin die aquatische Umwelt, während nicht-archosauriische Diapsiden weiterhin die Meeresumwelt dominierten. Auch die Temnospondyl- Linie großer Amphibien überlebte bis in die Kreidezeit in Australien (z. B. Koolasuchus ).
- Badlands in der Nähe von Drumheller , Alberta , wo Erosion die Kreide-Paläogen-Grenze freigelegt hat .Ereignis des Kreide-Paläogen-Aussterbens (Endkreidezeit, K-Pg-Aussterben oder früher K-T-Aussterben): 66 Ma , am Übergang von der Kreidezeit ( Maastricht ) zum Paläogen ( Danien ). [19] Das Ereignis wurde früher als Kreide-Tertiär- oder K-T-Aussterben oder K-T-Grenze bezeichnet; Es wird jetzt offiziell als Kreide-Paläogen-Aussterben (oder K-Pg-Aussterben) bezeichnet. Etwa 17 % aller Familien, 50 % aller Gattungen [6] und 75 % aller Arten starben aus. [2] In den Meeren verschwanden alle Ammoniten , Plesiosaurier und Mosasaurier und der Anteil sessiler Tiere sank auf etwa 33 %. Alle Nicht-Vogel- Dinosaurier starben in dieser Zeit aus. [20] Das Grenzereignis war schwerwiegend und zeigte erhebliche Schwankungen in der Aussterberate zwischen und zwischen verschiedenen Kladen . Säugetiere und Vögel , die ersteren von den Synapsiden und die letzteren von Theropodendinosauriern abstammen , entwickelten sich zu den dominanten Landtieren.
{kind=link}
{kind=link}
Trotz der Popularisierung dieser fünf Ereignisse gibt es keine eindeutige Grenze, die sie von anderen Aussterbeereignissen trennt; Die Verwendung unterschiedlicher Methoden zur Berechnung der Auswirkungen eines Aussterbens kann dazu führen, dass andere Ereignisse in den Top 5 aufgeführt werden. [21]
Ältere Fossilienfunde sind schwieriger zu interpretieren. Das ist weil:
- Ältere Fossilien sind schwerer zu finden, da sie meist in beträchtlicher Tiefe vergraben sind.
- Die Datierung älterer Fossilien ist schwieriger.
- Produktive Fossillagerstätten werden häufiger erforscht als unproduktive, sodass bestimmte Zeiträume unerforscht bleiben.
- Prähistorische Umweltereignisse können den Ablagerungsprozess stören .
- Die Erhaltung von Fossilien ist an Land unterschiedlich, Meeresfossilien sind jedoch tendenziell besser erhalten als ihre begehrteren Gegenstücke an Land. [22]
Es wurde vermutet, dass die scheinbaren Unterschiede in der marinen Biodiversität tatsächlich ein Artefakt sein könnten, wobei die Häufigkeitsschätzungen in direktem Zusammenhang mit der Menge an Gestein stehen, die für Proben aus verschiedenen Zeiträumen zur Verfügung steht. [23] Statistische Analysen zeigen jedoch, dass dies nur 50 % des beobachteten Musters erklären kann [ Zitat erforderlich ] und andere Beweise wie Pilzspitzen (geologisch schnelle Zunahme der Pilzhäufigkeit ) geben die Gewissheit, dass die meisten allgemein akzeptierten Aussterbeereignisse real sind . Eine Quantifizierung der Gesteinsexposition Westeuropas zeigt, dass viele der kleineren Ereignisse, für die eine biologische Erklärung gesucht wurde, am einfachsten durch Stichprobenverzerrungen erklärt werden können . [24]
Sechstes Massensterbenbearbeiten
Hauptartikel:
Verlust der biologischen Vielfalt
Untersuchungen, die nach der bahnbrechenden Veröffentlichung von 1982 (Sepkoski und Raup) abgeschlossen wurden, kamen zu dem Schluss, dass ein sechstes Massenaussterben aufgrund menschlicher Aktivitäten im Gange ist:
| 6. | Das Aussterben im Holozän ist derzeit im Gange. Seit 1900 ist die Aussterberate mehr als 1.000 Mal höher als die Hintergrundaussterberate , Tendenz steigend. [25] [26] [a] Das Massensterben ist eine Folge menschlicher Aktivitäten (ein Ökozid ) [28] [29] [30] [31] , angetrieben durch Bevölkerungswachstum und übermäßigen Verbrauch der natürlichen Ressourcen der Erde. [b] Die globale Biodiversitätsbewertung von IPBES aus dem Jahr 2019 besagt, dass von geschätzten 8 Millionen Arten derzeit 1 Million Pflanzen- und Tierarten vom Aussterben bedroht sind. [33] [34] [35] [36] Ende 2021 vermutete der WWF Deutschland, dass innerhalb eines Jahrzehnts über eine Million Arten aussterben könnten, was zum „größten Massenaussterben seit dem Ende des Dinosaurierzeitalters“ führen könnte. [37] Eine in PNAS veröffentlichte Studie aus dem Jahr 2023kam zu dem Schluss, dass seit 1500 mindestens 73 Tiergattungen ausgestorben sind. Hätte es den Menschen nie gegeben, hätte es 18.000 Jahre gedauert, bis dieselben Gattungen auf natürliche Weise verschwunden wären, heißt es in dem Bericht. [38] [39] [40] |
Aussterben nach Schweregradbearbeiten
Hauptartikel:
Aussterbeereignisse können mit verschiedenen Methoden verfolgt werden, darunter geologische Veränderungen, ökologische Auswirkungen, Aussterbe- vs. Entstehungsraten ( Artenbildung ) und am häufigsten der Diversitätsverlust zwischen taxonomischen Einheiten. Die meisten frühen Arbeiten verwendeten Familien als Einheit der Taxonomie, basierend auf Kompendien von Meerestierfamilien von Sepkoski (1982, 1992). [41] [42] Spätere Arbeiten von Sepkoski und anderen Autoren wechselten zu Gattungen , die präziser sind als Familien und weniger anfällig für taxonomische Verzerrungen oder unvollständige Stichproben im Vergleich zu Arten. [43] Hierbei handelt es sich um mehrere wichtige Veröffentlichungen, die den Verlust oder die ökologischen Auswirkungen von fünfzehn häufig diskutierten Aussterbeereignissen abschätzen. Im folgenden Abschnitt werden verschiedene Methoden beschrieben, die in diesen Arbeiten verwendet werden. Die „Big Five“-Massensterben sind fett gedruckt.
| Name des Aussterbens | Alter ( Ma ) | Sepkoski (1996) [44] Gattungen mit mehreren Intervallen | Bambach (2006) [45] | McGhee et al . (2013) [12] | Stanley (2016) [14] | |
|---|---|---|---|---|---|---|
| Taxonomischer Verlust | Ökologisches Ranking | |||||
| Spätes Ordovizium ( Ashgillian / Hirnantium ) | 445-444 | ~49 % | 57 % [d] (40 %, 31 %) [e] | 52 % | 7 | 42–46 % |
| Lau-Ereignis ( Ludfordian ) | 424 | ~23 % | – | 9 % | 9 | – |
| Kačák-Ereignis ( Eifelianisch ) | 388~ | ~24 % [a] | – | 32 % | 9 | – |
| Taghanisches Ereignis ( Givetian ) | 384~ | ~30 % [a] | 28,5 % | 36 % | 8 | – |
| Spätdevon / Kellwasser-Ereignis ( Frasnium ) | 372 | ~35 % | 34,7 % | 40 % | 4 | 16–20 % |
| End-Devon/ Hangenberg-Ereignis ( Famennium ) | 359 | ~28 % [a] | 31 % | 50 % | 7 | <13 % [w] |
| Serpukhovian | 330-325~ | ~23 % | 31 % | 39 % | 6 | 13-15 % |
| Kapitän | 260 | ~47 % [b] | 48 % | 25 % | 5 | 33-35 % |
| Perm–Trias ( Changhsingium ) | 252 | ~58 % | 55,7 % | 83 % | 1 | 62 % |
| Trias–Jura ( Rätium ) | 201 | ~37 % [c] | 47 % [c] | 73 % | 3 | N/A [g] |
| Pliensbachian-Toarcian | 186-178 | ~14 % | 25 %, 20 % [e] | – | – | – |
| Endjura (Tithonium) | 145 | ~18 % | 20 % | – | – | – |
| Cenomanisch-Turonisch | 94 | ~15 % | 25 % | – | – | – |
| Kreide-Paläogen ( Maastricht ) | 66 | ~39 % | 40–47 % | 40 % | 2 | 38–40 % |
| Eozän–Oligozän | 34 | ~11 % | 15,6 % | – | – | – |
a Von Sepkoski (1996) grafisch dargestellt, aber nicht diskutiert, wird als kontinuierlich mit dem Massenaussterben im späten Devon angesehen
b Zu der Zeit, die als kontinuierlich mit dem Massenaussterben am Ende des Perms betrachtet wird
c BeinhaltetspätnorianischeZeitabschnitte
d Diversitätsverlust beider Impulse zusammen berechnet
e Pulse erstrecken sich über benachbarte Zeitscheiben, separat berechnet
f Wird als ökologisch bedeutsam angesehen, aber nicht direkt analysiert
g Ausgeschlossen, da kein Konsens über die Chronologie der Spättrias besteht
Die Untersuchung großer Aussterbeereignissebearbeiten
Bahnbrechende Studien in den 1980er–1990er Jahrenbearbeiten

Während eines Großteils des 20. Jahrhunderts wurde die Untersuchung von Massenaussterben durch unzureichende Daten behindert. Obwohl Massenaussterben anerkannt wurden, galten sie als mysteriöse Ausnahmen von der vorherrschenden graduellen Sichtweise der Vorgeschichte, in der langsame Evolutionstendenzen die Veränderungen der Fauna bestimmen. Der erste Durchbruch wurde 1980 von einem Team unter der Leitung von Luis Alvarez veröffentlicht , das Spurenmetalle als Hinweise auf einen Asteroideneinschlag am Ende der Kreidezeit entdeckte . Die Alvarez-Hypothese für das Aussterben am Ende der Kreidezeit verschaffte Massenaussterben und katastrophalen Erklärungen neue öffentliche und wissenschaftliche Aufmerksamkeit. [46]

Eine weitere bahnbrechende Studie kam 1982, als ein Artikel von David M. Raup und Jack Sepkoski in der Zeitschrift Science veröffentlicht wurde . [2] In diesem Aufsatz, der auf einem von Sepkoski entwickelten Kompendium ausgestorbener Meerestierfamilien basiert , [ 41] wurden fünf Höhepunkte des Aussterbens von Meerestierfamilien identifiziert, die vor dem Hintergrund abnehmender Aussterberaten im Laufe der Zeit hervorstechen. Vier dieser Gipfel waren statistisch signifikant: das Aschgilium ( Endordovizium ), das Oberperm , das Norium ( Endtrias ) und das Maastrichtium (Endkreide). Der verbleibende Gipfel war ein breites Intervall mit hoher Auslöschung, das über die spätere Hälfte des Devon verschmiert war und dessen Höhepunkt im Frasnium- Stadium lag. [2]
In den 1980er Jahren haben Raup und Sepkoski ihre Aussterbe- und Entstehungsdaten weiter ausgearbeitet und darauf aufgebaut, indem sie eine hochauflösende Biodiversitätskurve (die „Sepkoski-Kurve“) und aufeinanderfolgende evolutionäre Faunen mit ihren eigenen Diversifizierungs- und Aussterbemustern definiert haben. [47] [48] [49] [50] [51] [52] Obwohl diese Interpretationen eine starke Grundlage für spätere Studien über Massenaussterben bildeten, schlugen Raup und Sepkoski 1984 auch eine kontroversere Idee vor: ein 26-Millionen-Jahres-Ereignis periodisches Muster bis hin zu Massenaussterben. [53] Zwei Astronomenteams brachten dies mit einem hypothetischen Braunen Zwerg in den entfernten Regionen des Sonnensystems in Verbindung und erfanden die „ Nemesis-Hypothese “, die von anderen Astronomen stark bestritten wurde .
Etwa zur gleichen Zeit begann Sepkoski mit der Ausarbeitung eines Kompendiums der Meerestiergattungen , das es Forschern ermöglichen sollte, das Aussterben mit einer feineren taxonomischen Auflösung zu untersuchen. Er begann bereits 1986 mit der Veröffentlichung vorläufiger Ergebnisse dieser laufenden Studie in einer Arbeit, in der 29 bemerkenswerte Aussterbeintervalle identifiziert wurden. [51] 1992 aktualisierte er auch sein Familienkompendium von 1982 und stellte fest, dass sich die Diversitätskurve trotz eines Jahrzehnts neuer Daten nur minimal veränderte. [42] [54] Im Jahr 1996 veröffentlichte Sepkoski eine weitere Arbeit, die das Aussterben mariner Gattungen (im Hinblick auf den Netto-Diversitätsverlust) nach Stadien verfolgte, ähnlich seiner früheren Arbeit über das Aussterben von Familien. Die Arbeit hat ihre Stichprobe auf drei Arten gefiltert: alle Gattungen (die gesamte ungefilterte Stichprobengröße), Gattungen mit mehreren Intervallen (nur solche, die in mehr als einem Stadium gefunden wurden) und „gut erhaltene“ Gattungen (ausgenommen solche aus Gruppen mit schlechter oder unzureichender Erforschung). Fossilienfunde). Basierend auf seinem Update von 1992 wurden auch Diversitätstrends in Meerestierfamilien überarbeitet. [44]
Das wiederbelebte Interesse an Massenaussterben veranlasste viele andere Autoren dazu, geologische Ereignisse im Kontext ihrer Auswirkungen auf das Leben neu zu bewerten. [55] In einer Arbeit von Michael Benton aus dem Jahr 1995 wurden die Aussterbe- und Entstehungsraten sowohl bei marinen als auch kontinentalen (Süßwasser- und Land-)Familien verfolgt und dabei 22 Aussterbeintervalle und kein periodisches Muster identifiziert. [56] Übersichtsbücher von OH Wallister (1996) und A. Hallam und PB Wignall (1997) fassen die neue Aussterbeforschung der letzten zwei Jahrzehnte zusammen. [57] [58] In einem Kapitel der früheren Quelle werden über 60 geologische Ereignisse aufgeführt, die möglicherweise als globale Aussterben unterschiedlicher Größenordnung angesehen werden könnten. [59] Diese Texte und andere weit verbreitete Veröffentlichungen in den 1990er Jahren trugen dazu bei, das populäre Bild von Massenaussterben als „Big Five“ neben vielen kleineren Aussterben in der Vorgeschichte zu etablieren.
Neue Daten zu Gattungen: Sepkoskis Kompendiumbearbeiten

Obwohl Sepkoski 1999 verstarb, wurde sein Kompendium über Meeresgattungen offiziell im Jahr 2002 veröffentlicht. Dies löste eine neue Welle von Studien zur Dynamik von Massenaussterben aus. [43] Diese Arbeiten nutzten das Kompendium, um die Entstehungsraten (die Rate, mit der neue Arten auftauchen oder sich bilden ) parallel zu den Aussterberaten im Kontext geologischer Stadien oder Unterstadien zu verfolgen. [60] Eine Überprüfung und erneute Analyse der Daten von Sepkoski durch Bambach (2006) identifizierte 18 verschiedene Massenaussterbeintervalle, darunter 4 große Aussterben im Kambrium . Diese entsprechen Sepkoskis Definition des Aussterbens, da es sich um kurze Unterstadien mit großem Diversitätsverlust und insgesamt hohen Aussterberaten im Vergleich zu ihrer Umgebung handelt. [45]
Bambach et al. (2004) gingen davon aus, dass jedes der „Big Five“-Aussterbeintervalle ein anderes Muster in der Beziehung zwischen Entstehung und Aussterbetrends aufweist. Darüber hinaus waren die Hintergrundaussterberaten weitgehend unterschiedlich und konnten in schwerwiegendere und weniger schwerwiegende Zeitintervalle unterteilt werden. Das Aussterben im Hintergrund war im Vergleich zur Entstehungsrate im mittleren Ordovizium/Untersilur, im späten Karbon/Perm und im jüngeren Jura am geringsten. Dies legt nahe, dass das Aussterben im späten Ordovizium, am Ende des Perms und am Ende der Kreidezeit statistisch signifikante Ausreißer in den Biodiversitätstrends waren, während das Aussterben im späten Devon und am Ende der Trias in Zeiträumen stattfand, die bereits durch ein relativ hohes Aussterben und eine geringe Entstehung belastet waren. [61]
Computermodelle von Foote (2005) ergaben, dass abrupte Aussterbeimpulse viel besser zum Muster der prähistorischen Artenvielfalt passen als eine allmähliche und kontinuierliche Hintergrundaussterberate mit sanften Spitzen und Tiefen. Dies unterstützt nachdrücklich den Nutzen schneller, häufiger Massenaussterben als Haupttreiber für Diversitätsveränderungen. Auch gepulste Ursprungsereignisse werden unterstützt, wenn auch in geringerem Maße, was weitgehend von gepulsten Auslöschungen abhängt. [62]
In ähnlicher Weise verwendete Stanley (2007) Aussterbe- und Entstehungsdaten, um Fluktuationsraten und Aussterbereaktionen verschiedener evolutionärer Faunen und taxonomischer Gruppen zu untersuchen. Im Gegensatz zu früheren Autoren deuten seine Diversitätssimulationen auf eine insgesamt exponentielle Wachstumsrate der Biodiversität im gesamten Phanerozoikum hin. [63]
Bekämpfung von Vorurteilen im Fossilienbestandbearbeiten

Als sich die Daten weiter anhäuften, begannen einige Autoren, Sepkoskis Stichprobe mit Methoden neu zu bewerten, die Stichprobenverzerrungen berücksichtigen sollten . Bereits 1982 stellten Phillip W. Signor und Jere H. Lipps in einem Artikel fest, dass die wahre Schärfe des Aussterbens durch die Unvollständigkeit des Fossilienbestands verwässert wurde. [64] Dieses Phänomen, später Signor-Lipps-Effekt genannt , besagt, dass das tatsächliche Aussterben einer Art nach ihrem letzten Fossil erfolgen muss und dass die Entstehung vor ihrem ersten Fossil erfolgen muss. Daher können Arten, die kurz vor einem plötzlichen Aussterben auszusterben scheinen, stattdessen Opfer des Ereignisses sein, obwohl allein anhand der Fossilienfunde ein offensichtlicher allmählicher Rückgang zu verzeichnen ist. Ein Modell von Foote (2007) ergab, dass in vielen geologischen Stadien die Aussterberaten künstlich erhöht wurden, weil Signor-Lipps spätere Stadien mit Aussterbeereignissen „zurückschmierte“. [65]

Zu den weiteren Verzerrungen gehört die Schwierigkeit, Taxa mit hohen Fluktuationsraten oder begrenzten Vorkommen zu beurteilen, die aufgrund mangelnder zeitlicher Feinauflösung nicht direkt beurteilt werden können. Um all diesen Verzerrungen Rechnung zu tragen, entscheiden sich viele Paläontologen dafür, Diversitätstrends durch zufällige Stichproben und die Verdünnung von Fossilienhäufigkeiten statt durch Rohdaten zur zeitlichen Reichweite zu beurteilen. Diese Lösung wird jedoch durch Vorurteile im Zusammenhang mit der Stichprobengröße beeinflusst. Eine große Voreingenommenheit ist insbesondere die „ Anziehungskraft des Neuen “, die Tatsache, dass sich der Fossilienbestand (und damit die bekannte Vielfalt) im Allgemeinen näher an der Moderne verbessert. Dies bedeutet, dass die Artenvielfalt und der Artenreichtum für ältere geologische Zeiträume möglicherweise allein aufgrund der Rohdaten unterschätzt werden. [60] [65] [3]
Alroy (2010) versuchte, stichprobenbedingte Verzerrungen bei Diversitätsschätzungen zu umgehen, indem er eine Methode verwendete, die er „ Shareholder Quorum Subsampling“ (SQS) nannte. Bei dieser Methode werden Fossilien aus einer „Sammlung“ (z. B. einem Zeitintervall) entnommen, um die relative Vielfalt dieser Sammlung zu beurteilen. Jedes Mal, wenn eine neue Art (oder ein anderes Taxon ) in die Probe gelangt, übernimmt sie alle anderen Fossilien dieser Art in die Sammlung (ihren „ Anteil “ an der Sammlung). Beispielsweise erreicht eine verzerrte Sammlung mit der Hälfte ihrer Fossilien von einer Art sofort einen Probenanteil von 50 %, wenn diese Art als erstes beprobt wird. Dies wird fortgesetzt, indem die Stichprobenanteile addiert werden, bis eine „Abdeckung“ oder ein „ Quorum “ erreicht ist, was sich auf eine voreingestellte gewünschte Summe von Anteilsprozentsätzen bezieht. Zu diesem Zeitpunkt wird die Anzahl der Arten in der Stichprobe gezählt. Es wird erwartet, dass eine Sammlung mit mehr Arten ein Stichprobenquorum mit mehr Arten erreicht und so die relative Diversitätsänderung zwischen zwei Sammlungen genau vergleicht, ohne sich auf die mit der Stichprobengröße verbundenen Verzerrungen zu verlassen. [66]
Alroy ging auch auf Drei-Timer-Algorithmen ein, die Verzerrungen bei Schätzungen der Aussterbe- und Entstehungsraten entgegenwirken sollen. Ein bestimmtes Taxon ist ein „Drei-Timer“, wenn es vor, nach und innerhalb eines bestimmten Zeitintervalls gefunden werden kann, und ein „Zwei-Timer“, wenn es sich auf einer Seite mit einem Zeitintervall überschneidet. Das Zählen von „Drei-Timern“ und „Zwei-Timern“ an beiden Enden eines Zeitintervalls und das Abtasten von Zeitintervallen nacheinander können zu Gleichungen kombiniert werden, um Aussterben und Entstehen mit weniger Verzerrungen vorherzusagen. [66] In späteren Arbeiten verfeinerte Alroy seine Gleichungen weiter, um bestehende Probleme mit Präzision und ungewöhnlichen Proben zu beheben. [67] [68]
McGhee et al. (2013), eine Arbeit, die sich hauptsächlich auf die ökologischen Auswirkungen von Massenaussterben konzentrierte, veröffentlichte ebenfalls neue Schätzungen der Schwere des Aussterbens auf der Grundlage von Alroys Methoden. Viele Aussterben waren nach diesen neuen Schätzungen deutlich schwerwiegender, einige waren jedoch weniger ausgeprägt. [12]
Stanley (2016) war ein weiterer Artikel, der versuchte, zwei häufige Fehler in früheren Schätzungen der Schwere des Aussterbens zu beseitigen. Der erste Fehler war die ungerechtfertigte Entfernung von „Singletons“, Gattungen, die nur in einem einzigen Zeitabschnitt vorkommen. Ihre Entfernung würde den Einfluss von Gruppen mit hohen Fluktuationsraten oder Abstammungslinien, die zu Beginn ihrer Diversifizierung unterbrochen wurden, verschleiern. Der zweite Fehler bestand darin, dass es schwierig war, Hintergrundaussterben von kurzen Massenaussterbeereignissen innerhalb desselben kurzen Zeitintervalls zu unterscheiden. Um dieses Problem zu umgehen, wurden die Hintergrundraten der Diversitätsveränderung (Aussterben/Entstehung) für Stadien oder Unterstadien ohne Massenaussterben geschätzt und dann davon ausgegangen, dass sie für nachfolgende Stadien mit Massenaussterben gelten. Beispielsweise wurden jeweils das Santonium- und das Campanium- Stadium verwendet, um Diversitätsänderungen im Maastrichtium vor dem K-Pg-Massenaussterben abzuschätzen. Das Abziehen der Hintergrundaussterben von den Aussterbezahlen hatte zur Folge, dass die geschätzte Schwere der sechs untersuchten Massensterbenereignisse verringert wurde. Dieser Effekt war bei Massenaussterben stärker, die in Perioden mit hohen Hintergrundaussterberaten auftraten, wie etwa im Devon. [14]
Unsicherheit im Proterozoikum und in früheren Äonenbearbeiten
Da der größte Teil der Vielfalt und Biomasse auf der Erde mikrobiell ist und sich daher anhand von Fossilien nur schwer messen lässt, betreffen die dokumentierten Aussterbeereignisse eher die leicht zu beobachtende, biologisch komplexe Komponente der Biosphäre als die gesamte Vielfalt und Fülle des Lebens. [69] Aus diesem Grund beschränken sich gut dokumentierte Aussterbeereignisse auf das Phanerozoikum – mit der einzigen Ausnahme der Sauerstoffkatastrophe im Proterozoikum –, da vor dem Phanerozoikum alle lebenden Organismen entweder mikrobiell oder, wenn mehrzellig, weichkörperig waren . Möglicherweise aufgrund des Fehlens eines robusten mikrobiellen Fossilienbestands scheinen Massenaussterben hauptsächlich ein Phänomen des Phanerozoikums zu sein, wobei lediglich die beobachtbaren Aussterberaten niedrig zu sein schienen, bevor große komplexe Organismen entstanden. [70]
Das Aussterben erfolgt mit ungleichmäßiger Geschwindigkeit. Basierend auf dem Fossilienbestand beträgt die Hintergrundrate des Aussterbens auf der Erde etwa zwei bis fünf taxonomische Familien von Meerestieren alle eine Million Jahre. Meeresfossilien werden aufgrund ihres überlegenen Fossilienbestands und ihrer stratigraphischen Reichweite im Vergleich zu Landtieren hauptsächlich zur Messung der Aussterberaten verwendet .
Die Sauerstoffkatastrophe, die sich vor rund 2,45 Milliarden Jahren im Paläoproterozoikum ereignete , gilt als das erste große Aussterben überhaupt. In gewisser Hinsicht war es vielleicht auch das Schlimmste, was es je gab, aber da die Ökologie der Erde kurz vor dieser Zeit so wenig verstanden wurde und das Konzept der Prokaryoten- Gattungen sich so sehr von den Gattungen komplexen Lebens unterschied, wäre es schwierig, es sinnvoll damit zu vergleichen keines der „Big Five“, auch wenn das Leben im Paläoproterozoikum besser bekannt wäre. [71]
Seit der kambrischen Explosion haben fünf weitere große Massenaussterben die Hintergrundaussterberate deutlich überschritten. Das jüngste und bekannteste Ereignis, das Kreide-Paläogen-Aussterben , das sich vor etwa 66 Ma (Millionen Jahren) ereignete, war ein groß angelegtes Massenaussterben von Tier- und Pflanzenarten in einem geologisch kurzen Zeitraum. [72] Zusätzlich zu den fünf großen Massensterben im Phanerozoikum gibt es zahlreiche kleinere, und das durch menschliche Aktivitäten verursachte anhaltende Massensterben wird manchmal als sechstes Massensterben bezeichnet . [73]
Evolutionäre Bedeutungbearbeiten
Siehe auch:
Massenaussterben haben manchmal die Entwicklung des Lebens auf der Erde beschleunigt . Wenn die Dominanz bestimmter ökologischer Nischen von einer Organismengruppe auf eine andere übergeht, liegt das selten daran, dass die neu dominierende Gruppe der alten „überlegen“ ist, sondern normalerweise daran, dass ein Aussterbeereignis die alte, dominante Gruppe vernichtet und Platz für die neue macht. ein Prozess, der als adaptive Strahlung bekannt ist . [74] [75]
Zum Beispiel existierten während der gesamten Herrschaft der Dinosaurier Säugetierformen („Fast-Säugetiere“) und dann Säugetiere , konnten aber in den großen Nischen der Landwirbeltiere, die die Dinosaurier monopolisierten, nicht konkurrieren. Das Massensterben am Ende der Kreidezeit vernichtete die Nichtvogeldinosaurier und ermöglichte es den Säugetieren, in die großen Nischen der Landwirbeltiere vorzudringen. Die Dinosaurier selbst waren Nutznießer eines früheren Massenaussterbens, des Endes der Trias , das die meisten ihrer Hauptkonkurrenten, die Crurotarsaner , vernichtet hatte .
Ein weiterer in der Eskalationshypothese vertretener Standpunkt geht davon aus, dass Arten in ökologischen Nischen mit mehr Konflikten zwischen Organismen eine geringere Wahrscheinlichkeit haben, das Aussterben zu überleben. Dies liegt daran, dass genau die Merkmale, die eine Art unter relativ statischen Bedingungen zahlreich und lebensfähig halten, zu einer Belastung werden, sobald die Populationszahlen konkurrierender Organismen während der Dynamik eines Aussterbeereignisses sinken.
Darüber hinaus erholen sich viele Gruppen, die ein Massenaussterben überleben, weder in ihrer Zahl noch in ihrer Vielfalt, und viele von ihnen verfallen langfristig, was oft als „ Dead Clades Walking “ bezeichnet wird. [76] Gruppen, die nach einem Massenaussterben noch längere Zeit überleben und auf nur wenige Arten reduziert wurden, dürften jedoch einen Rebound-Effekt erlebt haben, der als „ Push of the Past “ bezeichnet wird. [77]
Darwin war fest davon überzeugt, dass biotische Interaktionen wie der Wettbewerb um Nahrung und Raum – der „Kampf ums Dasein“ – für die Förderung von Evolution und Aussterben von wesentlich größerer Bedeutung seien als Veränderungen in der physischen Umwelt. Er drückte dies in „Der Ursprung der Arten“ aus :„Arten werden durch langsam wirkende Ursachen erzeugt und ausgerottet … und die wichtigste aller Ursachen organischer Veränderungen ist eine, die nahezu unabhängig von veränderten … physikalischen Bedingungen ist, nämlich die gegenseitige Beziehung von Organismus zu Organismus – die Verbesserung eines einzelnen Organismus, der die Verbesserung oder Ausrottung anderer nach sich zieht. [78]
https://translate.google.com/websitetranslationui?parent=https%3A%2F%2Fen-m-wikipedia-org.translate.goog&pfu=https%3A%2F%2Fen-m-wikipedia-org.translate.goog%2Fwiki%2FExtinction_event%3F_x_tr_sl%3Den%26_x_tr_tl%3Dde%26_x_tr_hl%3Dde%26_x_tr_pto%3Dwapp&u=https%3A%2F%2Fen.m.wikipedia.org%2Fwiki%2FExtinction_event&sl=en&tl=de&opu=https%3A%2F%2Ftranslate.google.com%2Fwebsite%3Fsl%3Den%26tl%3Dde%26hl%3Dde%26client%3Dwebapp%26u%3Dhttps%3A%2F%2Fen.m.wikipedia.org%2Fwiki%2FExtinction_event%26anno%3D2&client=webapp&hl=de
Aussterbeereignis
Zu diesem Thema sind auch von der Community erstellte Inhalte
- Automatische Ü
„Ereignis auf Extinktionsniveau“ leitet hier weiter. Für andere Verwendungen siehe
Extinction Level Event (Begriffsklärung) .In diesem Artikel geht es um das Massensterben. Für andere Verwendungen siehe
Extinction Event (Begriffsklärung) .
Ein Aussterben (auch bekannt als Massenaussterben oder biotische Krise ) ist ein weit verbreiteter und schneller Rückgang der Artenvielfalt auf der Erde . Ein solches Ereignis ist durch einen starken Rückgang der Vielfalt und Häufigkeit mehrzelliger Organismen gekennzeichnet . Es tritt auf, wenn die Aussterberate im Verhältnis zur Hintergrundaussterberate [1] und der Artbildungsrate zunimmt . Schätzungen zur Zahl der großen Massenaussterben in den letzten 540 Millionen Jahren reichen von nur fünf bis über zwanzig. Diese Unterschiede resultieren aus Uneinigkeit darüber, was ein „großes“ Aussterbeereignis darstellt und welche Daten zur Messung der vergangenen Vielfalt ausgewählt wurden.
Intensität des Meeressterbens während des Phanerozoikums
%
Vor Millionen von Jahren
Die blaue Grafik zeigt den scheinbaren
Prozentsatz (nicht die absolute Zahl) der Meerestiergattungen, die
in einem bestimmten Zeitraum ausgestorben sind. Es handelt sich nicht um alle Meeresarten, sondern nur um diejenigen, die leicht versteinerbar sind. Die Bezeichnungen der traditionellen „Big Five“-Aussterbeereignisse und des neueren bekannt gewordenen
Massenaussterbens auf den Capitanien sind anklickbare Links. Die beiden Aussterbeereignisse im Kambrium (ganz links) sind prozentual sehr groß, aber in absoluten Zahlen der bekannten Taxa gering, da zu dieser Zeit relativ wenig fossiles Leben produziert wurde.
Inhalt
Die „Big Five“-Massensterben
In einer bahnbrechenden Arbeit aus dem Jahr 1982 identifizierten Jack Sepkoski und David M. Raup fünf besondere geologische Abschnitte mit übermäßigem Diversitätsverlust. [2] Sie wurden ursprünglich als Ausreißer eines allgemeinen Trends sinkender Aussterberaten während des Phanerozoikums identifiziert, [3] aber als strengere statistische Tests auf die gesammelten Daten angewendet wurden, wurde festgestellt, dass im aktuellen Phanerozoikum Das vielzellige Tierleben hat mindestens fünf große und viele kleinere Massenaussterben erlebt. [4] Die „Big Five“ können nicht so klar definiert werden, sondern scheinen vielmehr die größten (oder einige der größten) eines relativ gleichmäßigen Kontinuums von Aussterbeereignissen darzustellen. [3] Es wird spekuliert, dass es sich um ein früheres (erstes?) Ereignis am Ende des Ediacaran handelte, und allen geht das vermutlich weitaus umfangreichere Massensterben mikrobiellen Lebens während der Sauerstoffkatastrophe zu Beginn des Proterozoikums voraus . [5]
- Ordovizium-Silur-Aussterbeereignisse (Endordovizium oder O–S): 445–444 Ma , kurz vor und am Ordovizium – Silur- Übergang. Es ereigneten sich zwei Ereignisse, die 27 % aller Familien , 57 % aller Gattungen und 85 % aller Arten töteten. [6] Zusammengenommen werden sie von vielen Wissenschaftlern als das zweitgrößte der fünf großen Artensterben in der Erdgeschichte eingestuft, gemessen am Prozentsatz derausgestorbenen Gattungen . Im Mai 2020 deuteten Studien darauf hin, dass die Ursachen des Massensterbens die globale Erwärmung im Zusammenhang mit Vulkanismus und Anoxie seien und nicht, wie zuvor angenommen, Abkühlung und Vereisung . [7] [8] Dies steht jedoch im Widerspruch zu zahlreichen früheren Studien, die die globale Abkühlung als Hauptursache angegeben haben. [9] In jüngster Zeit wurde vermutet, dass die Ablagerung von Vulkanasche der Auslöser für die Verringerung des atmosphärischen Kohlendioxids ist, was zu der in den geologischen Aufzeichnungen beobachteten Vereisung und Anoxie führt. [10]
- Aussterben im Spätdevon : 372–359 Ma , besetzt einen Großteil des Oberdevons bis zum Übergang Devon – Karbon . Das Oberdevon war ein Zeitraum mit hohem Diversitätsverlust, der sich auf zwei Aussterbeereignisse konzentrierte. Das größte Aussterben war das Kellwasser-Ereignis ( Frasnium – Famennium oder FF, 372 Ma), ein Aussterben am Ende des Frasniums, etwa in der Mitte des späten Devon. Dieses Aussterben vernichtete Korallenriffe und zahlreiche tropische benthische (auf dem Meeresboden lebende) Tiere wie kieferlose Fische, Brachiopoden und Trilobiten . Ein weiteres großes Aussterben war das Hangenberg-Ereignis (Devon-Karbon oder DC, 359 Ma), das das Ende des gesamten Devon bedeutete. Dieses Aussterben vernichtete die gepanzerten Placodermfische und führte beinahe zum Aussterben der neu entwickelten Ammonoide . Durch diese beiden kurz aufeinanderfolgenden Aussterbeereignisse wurden insgesamt etwa 19 % aller Familien, 50 % aller Gattungen [6] und mindestens 70 % aller Arten ausgelöscht. [11] Sepkoski und Raup (1982) [2] betrachteten das Aussterbenintervall im späten Devon ( Givetian- , Frasnium- und Famennium-Stadium) zunächst nicht als statistisch signifikant. [2] Unabhängig davon haben spätere Studien die starken ökologischen Auswirkungen der Kellwasser- und Hangenberg-Ereignisse bestätigt. [12]
- Trilobiten waren äußerst erfolgreiche Meerestiere, bis das Aussterben im Perm und der Trias sie alle auslöschte.Perm-Trias-Aussterbeereignis (Endperm): 252 Ma , am Perm – Trias- Übergang. [13] Das größte Aussterben des Phanerozoikums tötete 53 % der Meeresfamilien, 84 % der Meeresgattungen, etwa 81 % aller Meeresarten [14] und schätzungsweise 70 % der Landwirbeltierarten. [15] Dies ist auch das größte bekannte Aussterben von Insekten . [16] Der äußerst erfolgreiche Meeresgliederfüßer, der Trilobit , starb aus. Die Beweise für Pflanzen sind weniger eindeutig, aber nach dem Aussterben wurden neue Taxa dominant. [17] Das „Große Sterben“ hatte eine enorme evolutionäre Bedeutung: An Land beendete es die Vorherrschaft der frühen Synapsiden . Die Erholung der Wirbeltiere dauerte 30 Millionen Jahre, [18] aber die leeren Nischen boten den Archosauriern die Möglichkeit, auf dem Vormarsch zu. In den Meeren sank der Anteil der Tiere, die sich nicht fortbewegen konnten, von 67 % auf 50 %. Das gesamte späte Perm war eine schwierige Zeit, zumindest für das Meeresleben, sogar vor dem Aussterben der P-T-Grenze. Neuere Forschungen haben gezeigt, dass das End-Capitanian-Aussterben , das dem „Großen Sterben“ vorausging, wahrscheinlich ein vom P-T-Aussterben getrenntes Ereignis darstellt; Wenn ja, wäre es größer als einige der „Big Five“-Aussterbeereignisse und verdient möglicherweise einen separaten Platz in dieser Liste unmittelbar vor diesem.
- Trias-Jura-Aussterbeereignis (Endtrias): 201,3 Ma , am Übergang Trias – Jura . Etwa 23 % aller Familien, 48 % aller Gattungen (20 % der Meeresfamilien und 55 % der Meeresgattungen) und 70 bis 75 % aller Arten starben aus. [6] Die meisten nicht-dinosauriischen Archosaurier , die meisten Therapsiden und die meisten großen Amphibien wurden eliminiert, so dass die Dinosaurier nur noch wenig Konkurrenz auf der Erde hatten. Nicht-Dinosaurier-Archosaurier dominierten weiterhin die aquatische Umwelt, während nicht-archosauriische Diapsiden weiterhin die Meeresumwelt dominierten. Auch die Temnospondyl- Linie großer Amphibien überlebte bis in die Kreidezeit in Australien (z. B. Koolasuchus ).
- Badlands in der Nähe von Drumheller , Alberta , wo Erosion die Kreide-Paläogen-Grenze freigelegt hat .Ereignis des Kreide-Paläogen-Aussterbens (Endkreidezeit, K-Pg-Aussterben oder früher K-T-Aussterben): 66 Ma , am Übergang von der Kreidezeit ( Maastricht ) zum Paläogen ( Danien ). [19] Das Ereignis wurde früher als Kreide-Tertiär- oder K-T-Aussterben oder K-T-Grenze bezeichnet; Es wird jetzt offiziell als Kreide-Paläogen-Aussterben (oder K-Pg-Aussterben) bezeichnet. Etwa 17 % aller Familien, 50 % aller Gattungen [6] und 75 % aller Arten starben aus. [2] In den Meeren verschwanden alle Ammoniten , Plesiosaurier und Mosasaurier und der Anteil sessiler Tiere sank auf etwa 33 %. Alle Nicht-Vogel- Dinosaurier starben in dieser Zeit aus. [20] Das Grenzereignis war schwerwiegend und zeigte erhebliche Schwankungen in der Aussterberate zwischen und zwischen verschiedenen Kladen . Säugetiere und Vögel , die ersteren von den Synapsiden und die letzteren von Theropodendinosauriern abstammen , entwickelten sich zu den dominanten Landtieren.
Trotz der Popularisierung dieser fünf Ereignisse gibt es keine eindeutige Grenze, die sie von anderen Aussterbeereignissen trennt; Die Verwendung unterschiedlicher Methoden zur Berechnung der Auswirkungen eines Aussterbens kann dazu führen, dass andere Ereignisse in den Top 5 aufgeführt werden. [21]
Ältere Fossilienfunde sind schwieriger zu interpretieren. Das ist weil:
- Ältere Fossilien sind schwerer zu finden, da sie meist in beträchtlicher Tiefe vergraben sind.
- Die Datierung älterer Fossilien ist schwieriger.
- Produktive Fossillagerstätten werden häufiger erforscht als unproduktive, sodass bestimmte Zeiträume unerforscht bleiben.
- Prähistorische Umweltereignisse können den Ablagerungsprozess stören .
- Die Erhaltung von Fossilien ist an Land unterschiedlich, Meeresfossilien sind jedoch tendenziell besser erhalten als ihre begehrteren Gegenstücke an Land. [22]
Es wurde vermutet, dass die scheinbaren Unterschiede in der marinen Biodiversität tatsächlich ein Artefakt sein könnten, wobei die Häufigkeitsschätzungen in direktem Zusammenhang mit der Menge an Gestein stehen, die für Proben aus verschiedenen Zeiträumen zur Verfügung steht. [23] Statistische Analysen zeigen jedoch, dass dies nur 50 % des beobachteten Musters erklären kann [ Zitat erforderlich ] und andere Beweise wie Pilzspitzen (geologisch schnelle Zunahme der Pilzhäufigkeit ) geben die Gewissheit, dass die meisten allgemein akzeptierten Aussterbeereignisse real sind . Eine Quantifizierung der Gesteinsexposition Westeuropas zeigt, dass viele der kleineren Ereignisse, für die eine biologische Erklärung gesucht wurde, am einfachsten durch Stichprobenverzerrungen erklärt werden können . [24]
Sechstes Massensterben
Hauptartikel:
Verlust der biologischen Vielfalt
Untersuchungen, die nach der bahnbrechenden Veröffentlichung von 1982 (Sepkoski und Raup) abgeschlossen wurden, kamen zu dem Schluss, dass ein sechstes Massenaussterben aufgrund menschlicher Aktivitäten im Gange ist:
| 6. | Das Aussterben im Holozän ist derzeit im Gange. Seit 1900 ist die Aussterberate mehr als 1.000 Mal höher als die Hintergrundaussterberate , Tendenz steigend. [25] [26] [a] Das Massensterben ist eine Folge menschlicher Aktivitäten (ein Ökozid ) [28] [29] [30] [31] , angetrieben durch Bevölkerungswachstum und übermäßigen Verbrauch der natürlichen Ressourcen der Erde. [b] Die globale Biodiversitätsbewertung von IPBES aus dem Jahr 2019 besagt, dass von geschätzten 8 Millionen Arten derzeit 1 Million Pflanzen- und Tierarten vom Aussterben bedroht sind. [33] [34] [35] [36] Ende 2021 vermutete der WWF Deutschland, dass innerhalb eines Jahrzehnts über eine Million Arten aussterben könnten, was zum „größten Massenaussterben seit dem Ende des Dinosaurierzeitalters“ führen könnte. [37] Eine in PNAS veröffentlichte Studie aus dem Jahr 2023kam zu dem Schluss, dass seit 1500 mindestens 73 Tiergattungen ausgestorben sind. Hätte es den Menschen nie gegeben, hätte es 18.000 Jahre gedauert, bis dieselben Gattungen auf natürliche Weise verschwunden wären, heißt es in dem Bericht. [38] [39] [40] |
Aussterben nach Schweregrad
Hauptartikel:
Aussterbeereignisse können mit verschiedenen Methoden verfolgt werden, darunter geologische Veränderungen, ökologische Auswirkungen, Aussterbe- vs. Entstehungsraten ( Artenbildung ) und am häufigsten der Diversitätsverlust zwischen taxonomischen Einheiten. Die meisten frühen Arbeiten verwendeten Familien als Einheit der Taxonomie, basierend auf Kompendien von Meerestierfamilien von Sepkoski (1982, 1992). [41] [42] Spätere Arbeiten von Sepkoski und anderen Autoren wechselten zu Gattungen , die präziser sind als Familien und weniger anfällig für taxonomische Verzerrungen oder unvollständige Stichproben im Vergleich zu Arten. [43] Hierbei handelt es sich um mehrere wichtige Veröffentlichungen, die den Verlust oder die ökologischen Auswirkungen von fünfzehn häufig diskutierten Aussterbeereignissen abschätzen. Im folgenden Abschnitt werden verschiedene Methoden beschrieben, die in diesen Arbeiten verwendet werden. Die „Big Five“-Massensterben sind fett gedruckt.
| Name des Aussterbens | Alter ( Ma ) | Sepkoski (1996) [44] Gattungen mit mehreren Intervallen | Bambach (2006) [45] | McGhee et al . (2013) [12] | Stanley (2016) [14] | |
|---|---|---|---|---|---|---|
| Taxonomischer Verlust | Ökologisches Ranking | |||||
| Spätes Ordovizium ( Ashgillian / Hirnantium ) | 445-444 | ~49 % | 57 % [d] (40 %, 31 %) [e] | 52 % | 7 | 42–46 % |
| Lau-Ereignis ( Ludfordian ) | 424 | ~23 % | – | 9 % | 9 | – |
| Kačák-Ereignis ( Eifelianisch ) | 388~ | ~24 % [a] | – | 32 % | 9 | – |
| Taghanisches Ereignis ( Givetian ) | 384~ | ~30 % [a] | 28,5 % | 36 % | 8 | – |
| Spätdevon / Kellwasser-Ereignis ( Frasnium ) | 372 | ~35 % | 34,7 % | 40 % | 4 | 16–20 % |
| End-Devon/ Hangenberg-Ereignis ( Famennium ) | 359 | ~28 % [a] | 31 % | 50 % | 7 | <13 % [w] |
| Serpukhovian | 330-325~ | ~23 % | 31 % | 39 % | 6 | 13-15 % |
| Kapitän | 260 | ~47 % [b] | 48 % | 25 % | 5 | 33-35 % |
| Perm–Trias ( Changhsingium ) | 252 | ~58 % | 55,7 % | 83 % | 1 | 62 % |
| Trias–Jura ( Rätium ) | 201 | ~37 % [c] | 47 % [c] | 73 % | 3 | N/A [g] |
| Pliensbachian-Toarcian | 186-178 | ~14 % | 25 %, 20 % [e] | – | – | – |
| Endjura (Tithonium) | 145 | ~18 % | 20 % | – | – | – |
| Cenomanisch-Turonisch | 94 | ~15 % | 25 % | – | – | – |
| Kreide-Paläogen ( Maastricht ) | 66 | ~39 % | 40–47 % | 40 % | 2 | 38–40 % |
| Eozän–Oligozän | 34 | ~11 % | 15,6 % | – | – | – |
a Von Sepkoski (1996) grafisch dargestellt, aber nicht diskutiert, wird als kontinuierlich mit dem Massenaussterben im späten Devon angesehen
b Zu der Zeit, die als kontinuierlich mit dem Massenaussterben am Ende des Perms betrachtet wird
c BeinhaltetspätnorianischeZeitabschnitte
d Diversitätsverlust beider Impulse zusammen berechnet
e Pulse erstrecken sich über benachbarte Zeitscheiben, separat berechnet
f Wird als ökologisch bedeutsam angesehen, aber nicht direkt analysiert
g Ausgeschlossen, da kein Konsens über die Chronologie der Spättrias besteht
Die Untersuchung großer Aussterbeereignisse
Bahnbrechende Studien in den 1980er–1990er Jahren
Während eines Großteils des 20. Jahrhunderts wurde die Untersuchung von Massenaussterben durch unzureichende Daten behindert. Obwohl Massenaussterben anerkannt wurden, galten sie als mysteriöse Ausnahmen von der vorherrschenden graduellen Sichtweise der Vorgeschichte, in der langsame Evolutionstendenzen die Veränderungen der Fauna bestimmen. Der erste Durchbruch wurde 1980 von einem Team unter der Leitung von Luis Alvarez veröffentlicht , das Spurenmetalle als Hinweise auf einen Asteroideneinschlag am Ende der Kreidezeit entdeckte . Die Alvarez-Hypothese für das Aussterben am Ende der Kreidezeit verschaffte Massenaussterben und katastrophalen Erklärungen neue öffentliche und wissenschaftliche Aufmerksamkeit. [46]
Eine weitere bahnbrechende Studie kam 1982, als ein Artikel von David M. Raup und Jack Sepkoski in der Zeitschrift Science veröffentlicht wurde . [2] In diesem Aufsatz, der auf einem von Sepkoski entwickelten Kompendium ausgestorbener Meerestierfamilien basiert , [ 41] wurden fünf Höhepunkte des Aussterbens von Meerestierfamilien identifiziert, die vor dem Hintergrund abnehmender Aussterberaten im Laufe der Zeit hervorstechen. Vier dieser Gipfel waren statistisch signifikant: das Aschgilium ( Endordovizium ), das Oberperm , das Norium ( Endtrias ) und das Maastrichtium (Endkreide). Der verbleibende Gipfel war ein breites Intervall mit hoher Auslöschung, das über die spätere Hälfte des Devon verschmiert war und dessen Höhepunkt im Frasnium- Stadium lag. [2]
In den 1980er Jahren haben Raup und Sepkoski ihre Aussterbe- und Entstehungsdaten weiter ausgearbeitet und darauf aufgebaut, indem sie eine hochauflösende Biodiversitätskurve (die „Sepkoski-Kurve“) und aufeinanderfolgende evolutionäre Faunen mit ihren eigenen Diversifizierungs- und Aussterbemustern definiert haben. [47] [48] [49] [50] [51] [52] Obwohl diese Interpretationen eine starke Grundlage für spätere Studien über Massenaussterben bildeten, schlugen Raup und Sepkoski 1984 auch eine kontroversere Idee vor: ein 26-Millionen-Jahres-Ereignis periodisches Muster bis hin zu Massenaussterben. [53] Zwei Astronomenteams brachten dies mit einem hypothetischen Braunen Zwerg in den entfernten Regionen des Sonnensystems in Verbindung und erfanden die „ Nemesis-Hypothese “, die von anderen Astronomen stark bestritten wurde .
Etwa zur gleichen Zeit begann Sepkoski mit der Ausarbeitung eines Kompendiums der Meerestiergattungen , das es Forschern ermöglichen sollte, das Aussterben mit einer feineren taxonomischen Auflösung zu untersuchen. Er begann bereits 1986 mit der Veröffentlichung vorläufiger Ergebnisse dieser laufenden Studie in einer Arbeit, in der 29 bemerkenswerte Aussterbeintervalle identifiziert wurden. [51] 1992 aktualisierte er auch sein Familienkompendium von 1982 und stellte fest, dass sich die Diversitätskurve trotz eines Jahrzehnts neuer Daten nur minimal veränderte. [42] [54] Im Jahr 1996 veröffentlichte Sepkoski eine weitere Arbeit, die das Aussterben mariner Gattungen (im Hinblick auf den Netto-Diversitätsverlust) nach Stadien verfolgte, ähnlich seiner früheren Arbeit über das Aussterben von Familien. Die Arbeit hat ihre Stichprobe auf drei Arten gefiltert: alle Gattungen (die gesamte ungefilterte Stichprobengröße), Gattungen mit mehreren Intervallen (nur solche, die in mehr als einem Stadium gefunden wurden) und „gut erhaltene“ Gattungen (ausgenommen solche aus Gruppen mit schlechter oder unzureichender Erforschung). Fossilienfunde). Basierend auf seinem Update von 1992 wurden auch Diversitätstrends in Meerestierfamilien überarbeitet. [44]
Das wiederbelebte Interesse an Massenaussterben veranlasste viele andere Autoren dazu, geologische Ereignisse im Kontext ihrer Auswirkungen auf das Leben neu zu bewerten. [55] In einer Arbeit von Michael Benton aus dem Jahr 1995 wurden die Aussterbe- und Entstehungsraten sowohl bei marinen als auch kontinentalen (Süßwasser- und Land-)Familien verfolgt und dabei 22 Aussterbeintervalle und kein periodisches Muster identifiziert. [56] Übersichtsbücher von OH Wallister (1996) und A. Hallam und PB Wignall (1997) fassen die neue Aussterbeforschung der letzten zwei Jahrzehnte zusammen. [57] [58] In einem Kapitel der früheren Quelle werden über 60 geologische Ereignisse aufgeführt, die möglicherweise als globale Aussterben unterschiedlicher Größenordnung angesehen werden könnten. [59] Diese Texte und andere weit verbreitete Veröffentlichungen in den 1990er Jahren trugen dazu bei, das populäre Bild von Massenaussterben als „Big Five“ neben vielen kleineren Aussterben in der Vorgeschichte zu etablieren.
Neue Daten zu Gattungen: Sepkoskis Kompendium
Obwohl Sepkoski 1999 verstarb, wurde sein Kompendium über Meeresgattungen offiziell im Jahr 2002 veröffentlicht. Dies löste eine neue Welle von Studien zur Dynamik von Massenaussterben aus. [43] Diese Arbeiten nutzten das Kompendium, um die Entstehungsraten (die Rate, mit der neue Arten auftauchen oder sich bilden ) parallel zu den Aussterberaten im Kontext geologischer Stadien oder Unterstadien zu verfolgen. [60] Eine Überprüfung und erneute Analyse der Daten von Sepkoski durch Bambach (2006) identifizierte 18 verschiedene Massenaussterbeintervalle, darunter 4 große Aussterben im Kambrium . Diese entsprechen Sepkoskis Definition des Aussterbens, da es sich um kurze Unterstadien mit großem Diversitätsverlust und insgesamt hohen Aussterberaten im Vergleich zu ihrer Umgebung handelt. [45]
Bambach et al. (2004) gingen davon aus, dass jedes der „Big Five“-Aussterbeintervalle ein anderes Muster in der Beziehung zwischen Entstehung und Aussterbetrends aufweist. Darüber hinaus waren die Hintergrundaussterberaten weitgehend unterschiedlich und konnten in schwerwiegendere und weniger schwerwiegende Zeitintervalle unterteilt werden. Das Aussterben im Hintergrund war im Vergleich zur Entstehungsrate im mittleren Ordovizium/Untersilur, im späten Karbon/Perm und im jüngeren Jura am geringsten. Dies legt nahe, dass das Aussterben im späten Ordovizium, am Ende des Perms und am Ende der Kreidezeit statistisch signifikante Ausreißer in den Biodiversitätstrends waren, während das Aussterben im späten Devon und am Ende der Trias in Zeiträumen stattfand, die bereits durch ein relativ hohes Aussterben und eine geringe Entstehung belastet waren. [61]
Computermodelle von Foote (2005) ergaben, dass abrupte Aussterbeimpulse viel besser zum Muster der prähistorischen Artenvielfalt passen als eine allmähliche und kontinuierliche Hintergrundaussterberate mit sanften Spitzen und Tiefen. Dies unterstützt nachdrücklich den Nutzen schneller, häufiger Massenaussterben als Haupttreiber für Diversitätsveränderungen. Auch gepulste Ursprungsereignisse werden unterstützt, wenn auch in geringerem Maße, was weitgehend von gepulsten Auslöschungen abhängt. [62]
In ähnlicher Weise verwendete Stanley (2007) Aussterbe- und Entstehungsdaten, um Fluktuationsraten und Aussterbereaktionen verschiedener evolutionärer Faunen und taxonomischer Gruppen zu untersuchen. Im Gegensatz zu früheren Autoren deuten seine Diversitätssimulationen auf eine insgesamt exponentielle Wachstumsrate der Biodiversität im gesamten Phanerozoikum hin. [63]
Bekämpfung von Vorurteilen im Fossilienbestand
Als sich die Daten weiter anhäuften, begannen einige Autoren, Sepkoskis Stichprobe mit Methoden neu zu bewerten, die Stichprobenverzerrungen berücksichtigen sollten . Bereits 1982 stellten Phillip W. Signor und Jere H. Lipps in einem Artikel fest, dass die wahre Schärfe des Aussterbens durch die Unvollständigkeit des Fossilienbestands verwässert wurde. [64] Dieses Phänomen, später Signor-Lipps-Effekt genannt , besagt, dass das tatsächliche Aussterben einer Art nach ihrem letzten Fossil erfolgen muss und dass die Entstehung vor ihrem ersten Fossil erfolgen muss. Daher können Arten, die kurz vor einem plötzlichen Aussterben auszusterben scheinen, stattdessen Opfer des Ereignisses sein, obwohl allein anhand der Fossilienfunde ein offensichtlicher allmählicher Rückgang zu verzeichnen ist. Ein Modell von Foote (2007) ergab, dass in vielen geologischen Stadien die Aussterberaten künstlich erhöht wurden, weil Signor-Lipps spätere Stadien mit Aussterbeereignissen „zurückschmierte“. [65]
Zu den weiteren Verzerrungen gehört die Schwierigkeit, Taxa mit hohen Fluktuationsraten oder begrenzten Vorkommen zu beurteilen, die aufgrund mangelnder zeitlicher Feinauflösung nicht direkt beurteilt werden können. Um all diesen Verzerrungen Rechnung zu tragen, entscheiden sich viele Paläontologen dafür, Diversitätstrends durch zufällige Stichproben und die Verdünnung von Fossilienhäufigkeiten statt durch Rohdaten zur zeitlichen Reichweite zu beurteilen. Diese Lösung wird jedoch durch Vorurteile im Zusammenhang mit der Stichprobengröße beeinflusst. Eine große Voreingenommenheit ist insbesondere die „ Anziehungskraft des Neuen “, die Tatsache, dass sich der Fossilienbestand (und damit die bekannte Vielfalt) im Allgemeinen näher an der Moderne verbessert. Dies bedeutet, dass die Artenvielfalt und der Artenreichtum für ältere geologische Zeiträume möglicherweise allein aufgrund der Rohdaten unterschätzt werden. [60] [65] [3]
Alroy (2010) versuchte, stichprobenbedingte Verzerrungen bei Diversitätsschätzungen zu umgehen, indem er eine Methode verwendete, die er „ Shareholder Quorum Subsampling“ (SQS) nannte. Bei dieser Methode werden Fossilien aus einer „Sammlung“ (z. B. einem Zeitintervall) entnommen, um die relative Vielfalt dieser Sammlung zu beurteilen. Jedes Mal, wenn eine neue Art (oder ein anderes Taxon ) in die Probe gelangt, übernimmt sie alle anderen Fossilien dieser Art in die Sammlung (ihren „ Anteil “ an der Sammlung). Beispielsweise erreicht eine verzerrte Sammlung mit der Hälfte ihrer Fossilien von einer Art sofort einen Probenanteil von 50 %, wenn diese Art als erstes beprobt wird. Dies wird fortgesetzt, indem die Stichprobenanteile addiert werden, bis eine „Abdeckung“ oder ein „ Quorum “ erreicht ist, was sich auf eine voreingestellte gewünschte Summe von Anteilsprozentsätzen bezieht. Zu diesem Zeitpunkt wird die Anzahl der Arten in der Stichprobe gezählt. Es wird erwartet, dass eine Sammlung mit mehr Arten ein Stichprobenquorum mit mehr Arten erreicht und so die relative Diversitätsänderung zwischen zwei Sammlungen genau vergleicht, ohne sich auf die mit der Stichprobengröße verbundenen Verzerrungen zu verlassen. [66]
Alroy ging auch auf Drei-Timer-Algorithmen ein, die Verzerrungen bei Schätzungen der Aussterbe- und Entstehungsraten entgegenwirken sollen. Ein bestimmtes Taxon ist ein „Drei-Timer“, wenn es vor, nach und innerhalb eines bestimmten Zeitintervalls gefunden werden kann, und ein „Zwei-Timer“, wenn es sich auf einer Seite mit einem Zeitintervall überschneidet. Das Zählen von „Drei-Timern“ und „Zwei-Timern“ an beiden Enden eines Zeitintervalls und das Abtasten von Zeitintervallen nacheinander können zu Gleichungen kombiniert werden, um Aussterben und Entstehen mit weniger Verzerrungen vorherzusagen. [66] In späteren Arbeiten verfeinerte Alroy seine Gleichungen weiter, um bestehende Probleme mit Präzision und ungewöhnlichen Proben zu beheben. [67] [68]
McGhee et al. (2013), eine Arbeit, die sich hauptsächlich auf die ökologischen Auswirkungen von Massenaussterben konzentrierte, veröffentlichte ebenfalls neue Schätzungen der Schwere des Aussterbens auf der Grundlage von Alroys Methoden. Viele Aussterben waren nach diesen neuen Schätzungen deutlich schwerwiegender, einige waren jedoch weniger ausgeprägt. [12]
Stanley (2016) war ein weiterer Artikel, der versuchte, zwei häufige Fehler in früheren Schätzungen der Schwere des Aussterbens zu beseitigen. Der erste Fehler war die ungerechtfertigte Entfernung von „Singletons“, Gattungen, die nur in einem einzigen Zeitabschnitt vorkommen. Ihre Entfernung würde den Einfluss von Gruppen mit hohen Fluktuationsraten oder Abstammungslinien, die zu Beginn ihrer Diversifizierung unterbrochen wurden, verschleiern. Der zweite Fehler bestand darin, dass es schwierig war, Hintergrundaussterben von kurzen Massenaussterbeereignissen innerhalb desselben kurzen Zeitintervalls zu unterscheiden. Um dieses Problem zu umgehen, wurden die Hintergrundraten der Diversitätsveränderung (Aussterben/Entstehung) für Stadien oder Unterstadien ohne Massenaussterben geschätzt und dann davon ausgegangen, dass sie für nachfolgende Stadien mit Massenaussterben gelten. Beispielsweise wurden jeweils das Santonium- und das Campanium- Stadium verwendet, um Diversitätsänderungen im Maastrichtium vor dem K-Pg-Massenaussterben abzuschätzen. Das Abziehen der Hintergrundaussterben von den Aussterbezahlen hatte zur Folge, dass die geschätzte Schwere der sechs untersuchten Massensterbenereignisse verringert wurde. Dieser Effekt war bei Massenaussterben stärker, die in Perioden mit hohen Hintergrundaussterberaten auftraten, wie etwa im Devon. [14]
Unsicherheit im Proterozoikum und in früheren Äonen
Da der größte Teil der Vielfalt und Biomasse auf der Erde mikrobiell ist und sich daher anhand von Fossilien nur schwer messen lässt, betreffen die dokumentierten Aussterbeereignisse eher die leicht zu beobachtende, biologisch komplexe Komponente der Biosphäre als die gesamte Vielfalt und Fülle des Lebens. [69] Aus diesem Grund beschränken sich gut dokumentierte Aussterbeereignisse auf das Phanerozoikum – mit der einzigen Ausnahme der Sauerstoffkatastrophe im Proterozoikum –, da vor dem Phanerozoikum alle lebenden Organismen entweder mikrobiell oder, wenn mehrzellig, weichkörperig waren . Möglicherweise aufgrund des Fehlens eines robusten mikrobiellen Fossilienbestands scheinen Massenaussterben hauptsächlich ein Phänomen des Phanerozoikums zu sein, wobei lediglich die beobachtbaren Aussterberaten niedrig zu sein schienen, bevor große komplexe Organismen entstanden. [70]
Das Aussterben erfolgt mit ungleichmäßiger Geschwindigkeit. Basierend auf dem Fossilienbestand beträgt die Hintergrundrate des Aussterbens auf der Erde etwa zwei bis fünf taxonomische Familien von Meerestieren alle eine Million Jahre. Meeresfossilien werden aufgrund ihres überlegenen Fossilienbestands und ihrer stratigraphischen Reichweite im Vergleich zu Landtieren hauptsächlich zur Messung der Aussterberaten verwendet .
Die Sauerstoffkatastrophe, die sich vor rund 2,45 Milliarden Jahren im Paläoproterozoikum ereignete , gilt als das erste große Aussterben überhaupt. In gewisser Hinsicht war es vielleicht auch das Schlimmste, was es je gab, aber da die Ökologie der Erde kurz vor dieser Zeit so wenig verstanden wurde und das Konzept der Prokaryoten- Gattungen sich so sehr von den Gattungen komplexen Lebens unterschied, wäre es schwierig, es sinnvoll damit zu vergleichen keines der „Big Five“, auch wenn das Leben im Paläoproterozoikum besser bekannt wäre. [71]
Seit der kambrischen Explosion haben fünf weitere große Massenaussterben die Hintergrundaussterberate deutlich überschritten. Das jüngste und bekannteste Ereignis, das Kreide-Paläogen-Aussterben , das sich vor etwa 66 Ma (Millionen Jahren) ereignete, war ein groß angelegtes Massenaussterben von Tier- und Pflanzenarten in einem geologisch kurzen Zeitraum. [72] Zusätzlich zu den fünf großen Massensterben im Phanerozoikum gibt es zahlreiche kleinere, und das durch menschliche Aktivitäten verursachte anhaltende Massensterben wird manchmal als sechstes Massensterben bezeichnet . [73]
Evolutionäre Bedeutung
Siehe auch:
Massenaussterben haben manchmal die Entwicklung des Lebens auf der Erde beschleunigt . Wenn die Dominanz bestimmter ökologischer Nischen von einer Organismengruppe auf eine andere übergeht, liegt das selten daran, dass die neu dominierende Gruppe der alten „überlegen“ ist, sondern normalerweise daran, dass ein Aussterbeereignis die alte, dominante Gruppe vernichtet und Platz für die neue macht. ein Prozess, der als adaptive Strahlung bekannt ist . [74] [75]
Zum Beispiel existierten während der gesamten Herrschaft der Dinosaurier Säugetierformen („Fast-Säugetiere“) und dann Säugetiere , konnten aber in den großen Nischen der Landwirbeltiere, die die Dinosaurier monopolisierten, nicht konkurrieren. Das Massensterben am Ende der Kreidezeit vernichtete die Nichtvogeldinosaurier und ermöglichte es den Säugetieren, in die großen Nischen der Landwirbeltiere vorzudringen. Die Dinosaurier selbst waren Nutznießer eines früheren Massenaussterbens, des Endes der Trias , das die meisten ihrer Hauptkonkurrenten, die Crurotarsaner , vernichtet hatte .
Ein weiterer in der Eskalationshypothese vertretener Standpunkt geht davon aus, dass Arten in ökologischen Nischen mit mehr Konflikten zwischen Organismen eine geringere Wahrscheinlichkeit haben, das Aussterben zu überleben. Dies liegt daran, dass genau die Merkmale, die eine Art unter relativ statischen Bedingungen zahlreich und lebensfähig halten, zu einer Belastung werden, sobald die Populationszahlen konkurrierender Organismen während der Dynamik eines Aussterbeereignisses sinken.
Darüber hinaus erholen sich viele Gruppen, die ein Massenaussterben überleben, weder in ihrer Zahl noch in ihrer Vielfalt, und viele von ihnen verfallen langfristig, was oft als „ Dead Clades Walking “ bezeichnet wird. [76] Gruppen, die nach einem Massenaussterben noch längere Zeit überleben und auf nur wenige Arten reduziert wurden, dürften jedoch einen Rebound-Effekt erlebt haben, der als „ Push of the Past “ bezeichnet wird. [77]
Darwin war fest davon überzeugt, dass biotische Interaktionen wie der Wettbewerb um Nahrung und Raum – der „Kampf ums Dasein“ – für die Förderung von Evolution und Aussterben von wesentlich größerer Bedeutung seien als Veränderungen in der physischen Umwelt. Er drückte dies in „Der Ursprung der Arten“ aus :„Arten werden durch langsam wirkende Ursachen erzeugt und ausgerottet … und die wichtigste aller Ursachen organischer Veränderungen ist eine, die nahezu unabhängig von veränderten … physikalischen Bedingungen ist, nämlich die gegenseitige Beziehung von Organismus zu Organismus – die Verbesserung eines einzelnen Organismus, der die Verbesserung oder Ausrottung anderer nach sich zieht. [78]
Muster in der Häufigkeit
Verschiedene Autoren haben vorgeschlagen, dass Aussterbeereignisse regelmäßig alle 26 bis 30 Millionen Jahre auftraten [79] [53] oder dass die Vielfalt etwa alle 62 Millionen Jahre episodisch schwankt. [80] Verschiedene Ideen, meist in Bezug auf astronomische Einflüsse, versuchen, das vermutete Muster zu erklären, darunter die Anwesenheit eines hypothetischen Begleitsterns zur Sonne, [81] [82] Schwingungen in der galaktischen Ebene oder der Durchgang durch die Spiralarme der Milchstraße . [83] Andere Autoren sind jedoch zu dem Schluss gekommen, dass die Daten zu Massenaussterben im Meer nicht mit der Vorstellung übereinstimmen, dass Massenaussterben periodisch sind oder dass sich Ökosysteme allmählich bis zu einem Punkt aufbauen, an dem ein Massenaussterben unvermeidlich ist. [3] Viele der vorgeschlagenen Korrelationen wurden als falsch oder ohne statistische Signifikanz angesehen. [84] [85] [86] Andere haben argumentiert, dass es starke Beweise für die Periodizität in einer Vielzahl von Aufzeichnungen gibt, [87] und zusätzliche Beweise in Form einer zusammenfallenden periodischen Variation in nichtbiologischen geochemischen Variablen wie Strontiumisotopen, [88] Überschwemmungsbasalte, anoxische Ereignisse, Orogenese und Evaporitablagerung. Eine Erklärung für diesen vorgeschlagenen Zyklus ist die Speicherung und Freisetzung von Kohlenstoff durch die ozeanische Kruste, die Kohlenstoff zwischen Atmosphäre und Mantel austauscht. [89]
Alle Gattungen
„Gut definierte“ Gattungen
Trendlinie
„Big Five“ -Massensterben
Andere Massensterben
Vor Millionen Jahren
Tausende Gattungen
Phanerozoische Artenvielfalt, wie der Fossilienbestand zeigt
Es wird angenommen, dass es zu Massenaussterben kommt, wenn ein langfristiger Stress durch einen kurzfristigen Schock verstärkt wird. [90] Im Laufe des Phanerozoikums scheint die Wahrscheinlichkeit des Aussterbens einzelner Taxa geringer geworden zu sein, [91] was auf robustere Nahrungsnetze sowie weniger vom Aussterben bedrohte Arten und andere Faktoren wie die kontinentale Verbreitung zurückzuführen sein könnte. [91] Selbst nach Berücksichtigung der Stichprobenverzerrung scheint es jedoch während des Phanerozoikums zu einem allmählichen Rückgang der Aussterbe- und Entstehungsraten zu kommen. [3] Dies könnte auf die Tatsache zurückzuführen sein, dass Gruppen mit höheren Fluktuationsraten eher durch Zufall aussterben; Oder es kann ein Artefakt der Taxonomie sein: Familien neigen dazu, mit der Zeit artenreicher zu werden und daher weniger anfällig für das Aussterben zu sein. [3] und größere taxonomische Gruppen (per Definition) erscheinen früher in der geologischen Zeit. [92]
Es wurde auch vermutet, dass die Ozeane im Laufe der letzten 500 Millionen Jahre nach und nach lebensfreundlicher geworden sind und daher weniger anfällig für Massenaussterben [c] [93] [94] , eine Anfälligkeit für Aussterben auf taxonomischer Ebene scheint jedoch nicht erkennbar Massenaussterben mehr oder weniger wahrscheinlich zu machen. [91]
Ursachen
Es gibt immer noch Debatten über die Ursachen aller Massensterben. Im Allgemeinen kann es zu großen Artensterben kommen, wenn eine Biosphäre unter Langzeitstress einen kurzfristigen Schock erleidet. [90] Ein zugrunde liegender Mechanismus scheint in der Korrelation der Aussterbe- und Entstehungsraten mit der Diversität zu liegen. Eine hohe Diversität führt zu einem anhaltenden Anstieg der Aussterberate; geringe Diversität führt zu einem anhaltenden Anstieg der Entstehungsrate. Diese vermutlich ökologisch kontrollierten Beziehungen verstärken wahrscheinlich kleinere Störungen (Asteroideneinschläge usw.), um die beobachteten globalen Auswirkungen hervorzurufen. [3]
Ursachen für bestimmte Massensterben identifizieren
Eine gute Theorie für ein bestimmtes Massensterben sollte:
- Erklären Sie alle Verluste und konzentrieren Sie sich nicht nur auf einige wenige Gruppen (z. B. Dinosaurier).
- erklären, warum bestimmte Gruppen von Organismen ausgestorben sind und warum andere überlebt haben;
- Bereitstellung von Mechanismen, die stark genug sind, um ein Massensterben, aber kein vollständiges Aussterben auszulösen;
- auf Ereignissen oder Prozessen basieren, die nachweislich stattgefunden haben, und nicht nur auf dem Aussterben.
Möglicherweise müssen Kombinationen von Ursachen berücksichtigt werden. Beispielsweise scheint der marine Aspekt des Aussterbens am Ende der Kreidezeit durch mehrere Prozesse verursacht worden zu sein, die sich zeitlich teilweise überschnitten und möglicherweise in verschiedenen Teilen der Welt unterschiedliche Bedeutung hatten. [95]
Arens und West (2006) schlugen ein „Presse/Pulse“-Modell vor, bei dem Massenaussterben im Allgemeinen zwei Arten von Ursachen erfordern: langfristigen Druck auf das Ökosystem („Presse“) und eine plötzliche Katastrophe („Pulse“) in Richtung des Ende der Druckperiode. [96] Ihre statistische Analyse der marinen Aussterberaten im gesamten Phanerozoikum legte nahe, dass weder ein langfristiger Druck allein noch eine Katastrophe allein ausreichten, um einen signifikanten Anstieg der Aussterberate zu verursachen.
Die am weitesten verbreiteten Erklärungen
MacLeod (2001) [97] fasste den Zusammenhang zwischen Massenaussterben und Ereignissen zusammen, die am häufigsten als Ursachen für Massenaussterben genannt werden, und verwendete dabei Daten von Courtillot, Jaeger & Yang et al. (1996), [98] Hallam (1992) [99] und Grieve & Pesonen (1992): [100]
- Überschwemmungs-Basalt- Ereignisse (riesige Vulkanausbrüche): 11 Vorkommnisse, alle mit bedeutenden Aussterben verbunden [d] [e] Wignall (2001) kam jedoch zu dem Schluss, dass nur fünf der großen Aussterben mit Überschwemmungs-Basalteruptionen zusammenfielen und dass die Hauptphase des Aussterbens vorher begann die Eruptionen. [101]
- Meeresspiegelrückgänge: 12, davon waren sieben mit bedeutenden Artensterben verbunden. [e]
- Asteroideneinschläge : Ein großer Einschlag ist mit einem Massenaussterben verbunden, d. h. dem Kreide-Paläogen-Aussterben. Es gab viele kleinere Einschläge, die jedoch nicht mit bedeutenden Artensterben in Verbindung gebracht wurden [102] oder nicht genau genug datiert werden konnten. Der Einschlag, der den Siljan-Ring hervorbrachte, erfolgte entweder kurz vor dem Aussterben im späten Devon oder fiel damit zusammen. [103]
Die am häufigsten vermuteten Ursachen für Massensterben sind unten aufgeführt.
Überschwemmungsbasaltereignisse

Die Bildung großer magmatischer Provinzen durch Überschwemmungsbasaltereignisse könnte Folgendes zur Folge haben:
- produzierte Staub und Partikelaerosole , die die Photosynthese hemmten und so zum Zusammenbruch der Nahrungsketten sowohl an Land als auch auf See führten [104]
- emittierte Schwefeloxide, die sich als saurer Regen niederschlugen , viele Organismen vergifteten und so zum Zusammenbruch der Nahrungsketten beitrugen
- Es emittierte Kohlendioxid und verursachte möglicherweise eine nachhaltige globale Erwärmung, sobald sich der Staub und die Partikelaerosole verflüchtigten.
Basaltflutereignisse treten als Aktivitätsimpulse auf, die von Ruhephasen unterbrochen werden. Infolgedessen dürften sie dazu führen, dass das Klima zwischen Abkühlung und Erwärmung schwankt, allerdings mit einem allgemeinen Trend zur Erwärmung, da das von ihnen emittierte Kohlendioxid Hunderte von Jahren in der Atmosphäre verbleiben kann.
Basaltüberschwemmungen gelten als Ursache für viele große Artensterben. [105] [106] Es wird spekuliert, dass massiver Vulkanismus das Kellwasser-Ereignis , [107] [108] [109] das End-Guadalupian-Aussterben und [110] [111] [112] das End-Perm-Aussterben verursacht oder dazu beigetragen hat Ereignis , [113] [114] [115] das Smithian-Spathian-Aussterben , [116] [117] [118] das Trias-Jura-Aussterben , [119] [120] [121] das toarcische ozeanische anoxische Ereignis , [122 ] [123] [124] das ozeanische anoxische Ereignis im Cenoman-Turon , [125] [126] [127] das Kreide-Paläogen-Aussterben , [128] [129] [130] und das paläozän-eozäne thermische Maximum . [131] [132] [133] Der Zusammenhang zwischen gigantischen Vulkanereignissen, die sich in den großen magmatischen Provinzen äußerten, und Massenaussterben wurde für die letzten 260 Millionen Jahre gezeigt. [134] [135] Kürzlich wurde eine solche mögliche Korrelation auf das gesamte Phanerozoikum ausgedehnt . [136]
Rückgang des Meeresspiegels
Diese sind oft deutlich durch weltweite Abfolgen zeitgenössischer Sedimente gekennzeichnet, die den Übergang vom Meeresboden zur Gezeitenzone zum Strand zum Festland ganz oder teilweise zeigen – und bei denen es keine Hinweise darauf gibt, dass die Gesteine in den relevanten Gebieten durch geologische Prozesse angehoben wurden wie zum Beispiel die Orogenese . Ein Rückgang des Meeresspiegels könnte die Fläche des Festlandsockels (den produktivsten Teil der Ozeane) soweit verkleinern, dass es zu einem Massensterben im Meer kommt, und könnte die Wetterverhältnisse so weit stören, dass es zu Aussterben an Land kommt. Der Rückgang des Meeresspiegels ist jedoch sehr wahrscheinlich das Ergebnis anderer Ereignisse, etwa einer anhaltenden globalen Abkühlung oder des Absinkens der mittelozeanischen Rücken .
Der Rückgang des Meeresspiegels wird mit den meisten Massenaussterben in Verbindung gebracht, einschließlich aller „Big Five“ – Endordovizium , Oberdevon , Endperm , Endtrias und Endkreide sowie das erst kürzlich erkannte Kapitanische Massenaussterben Aussterben von vergleichbarer Schwere wie die Big Five. [137] [138]
Eine 2008 in der Fachzeitschrift Nature veröffentlichte Studie stellte einen Zusammenhang zwischen der Geschwindigkeit von Massensterben und Veränderungen des Meeresspiegels und der Sedimente her. [139] Die Studie legt nahe, dass Veränderungen in der Meeresumgebung im Zusammenhang mit dem Meeresspiegel einen treibenden Einfluss auf die Aussterberaten haben und im Allgemeinen die Zusammensetzung des Lebens in den Ozeanen bestimmen. [140]
Außerirdische Bedrohungen
Impact-Ereignisse

Der Einschlag eines ausreichend großen Asteroiden oder Kometen hätte sowohl an Land als auch auf See zum Zusammenbruch der Nahrungsketten führen können , indem er Staub und Partikelaerosole erzeugte und so die Photosynthese hemmte. [141] Einschläge auf schwefelreiches Gestein könnten Schwefeloxide freigesetzt haben, die als giftiger saurer Regen ausfielen und so zum Zusammenbruch der Nahrungsketten beitrugen. Solche Einschläge könnten auch zu Megatsunamis und/oder globalen Waldbränden geführt haben .
Die meisten Paläontologen sind sich inzwischen einig, dass vor etwa 66 Millionen Jahren ein Asteroid die Erde getroffen hat, es besteht jedoch weiterhin Streit darüber, ob der Einschlag die einzige Ursache für das Kreide-Paläogen-Aussterben war . [142] [143] Dennoch berichteten Forscher im Oktober 2019, dass der Asteroideneinschlag von Chicxulub in der Kreidezeit , der vor 66 Millionen Jahren zum Aussterben von Nicht-Vogel -Dinosauriern führte, auch die Ozeane schnell versaute , was zu einem ökologischen Zusammenbruch und langanhaltenden Auswirkungen auf das Klima führte und war einer der Hauptgründe für das Massensterben am Ende der Kreidezeit. [144] [145]
Es wurde auch die Hypothese aufgestellt, dass das Aussterben im Perm-Trias durch einen Asteroideneinschlag verursacht wurde, der den Araguainha-Krater bildete , da sich das geschätzte Datum der Kraterbildung mit dem Aussterben am Ende des Perms überschnitt. [146] [147] [148] Diese Hypothese wurde jedoch weitgehend in Frage gestellt, wobei die Auswirkungshypothese von den meisten Forschern abgelehnt wurde. [149] [150] [151]
Nach der Shiva-Hypothese ist die Erde aufgrund des Durchgangs der Sonne durch die Ebene der Milchstraße etwa alle 27 Millionen Jahre verstärkten Asteroideneinschlägen ausgesetzt , was alle 27 Millionen Jahre zu Aussterbeereignissen führt. Einige Beweise für diese Hypothese wurden sowohl im marinen als auch im nichtmarinen Kontext gefunden. [152] Alternativ könnte der Durchgang der Sonne durch die Spiralarme der Galaxie mit höherer Dichte mit einem Massenaussterben auf der Erde zusammenfallen, möglicherweise aufgrund häufigerer Einschlagereignisse . [153] Eine erneute Analyse der Auswirkungen des Sonnendurchgangs durch die Spiralstruktur auf der Grundlage von Karten der Spiralstruktur der Milchstraße auf die CO-Moleküllinienemission konnte jedoch keinen Zusammenhang finden. [154]
Eine nahegelegene Nova, Supernova oder ein Gammastrahlenausbruch
Ein nahegelegener Gammastrahlenausbruch (weniger als 6000 Lichtjahre entfernt) wäre stark genug, um die Ozonschicht der Erde zu zerstören und Organismen für die ultraviolette Strahlung der Sonne anfällig zu machen. [155] Gammastrahlenausbrüche sind ziemlich selten und treten in einer bestimmten Galaxie nur wenige Male pro Million Jahre auf. [156] Es wurde vermutet, dass ein Gammastrahlenausbruch das Aussterben im Endordovizium verursachte , [157] [158] während eine Supernova als Ursache für das Hangenberg-Ereignis vorgeschlagen wurde . [159]
Globale Abkühlung
Eine anhaltende und erhebliche globale Abkühlung könnte viele polare und gemäßigte Arten töten und andere dazu zwingen, in Richtung Äquator zu wandern . die für tropische Arten verfügbare Fläche reduzieren ; führen oft dazu, dass das Klima auf der Erde im Durchschnitt trockener wird, vor allem dadurch, dass mehr Wasser auf dem Planeten in Eis und Schnee eingeschlossen wird. Man geht davon aus, dass die Vereisungszyklen der aktuellen Eiszeit nur sehr geringe Auswirkungen auf die Artenvielfalt hatten, so dass die bloße Existenz einer deutlichen Abkühlung allein nicht ausreicht, um ein Massensterben zu erklären.
Es wurde vermutet, dass die globale Abkühlung das Aussterben im Endordovizium , im Perm-Trias , im späten Devon und möglicherweise auch in anderen Ausbrüchen verursachte oder dazu beitrug. Eine nachhaltige globale Abkühlung unterscheidet sich von den vorübergehenden klimatischen Auswirkungen von Hochwasserbasaltereignissen oder -einschlägen.
Globale Erwärmung
Hauptartikel:
Aussterberisiko durch globale Erwärmung
Dies hätte den gegenteiligen Effekt: die für tropische Arten verfügbare Fläche würde vergrößert; Töten Sie gemäßigte Arten oder zwingen Sie sie, in Richtung der Pole zu wandern ; möglicherweise schweres Aussterben polarer Arten verursachen; führen oft dazu, dass das Klima auf der Erde im Durchschnitt feuchter wird, vor allem indem sie Eis und Schnee schmelzen und so das Volumen des Wasserkreislaufs vergrößern . Es könnte auch zu anoxischen Ereignissen in den Ozeanen führen (siehe unten).
Mehrere neuere Studien belegen, dass die globale Erwärmung eine Ursache für das Massensterben ist. [160]
Das dramatischste Beispiel einer anhaltenden Erwärmung ist das paläozän-eozäne thermische Maximum , das mit einem der kleineren Massenaussterben verbunden war. Es wurde auch vermutet, dass es das Trias-Jura-Aussterbeereignis verursacht hat , bei dem 20 % aller Meeresfamilien ausstarben. Darüber hinaus wurde vermutet, dass das Aussterben im Perm-Trias durch die Erwärmung verursacht wurde. [161] [162] [163]
Clathrat-Waffen-Hypothese
Hauptartikel:
Clathrate sind Verbundstoffe, bei denen ein Gitter einer Substanz einen Käfig um eine andere bildet. Auf Festlandsockeln bilden sich Methanclathrate (in denen Wassermoleküle den Käfig bilden) . Diese Clathrate zerfallen wahrscheinlich schnell und geben das Methan frei, wenn die Temperatur schnell ansteigt oder der Druck auf sie schnell abfällt – beispielsweise als Reaktion auf eine plötzliche globale Erwärmung oder einen plötzlichen Rückgang des Meeresspiegels oder sogar Erdbeben . Methan ist ein viel stärkeres Treibhausgas als Kohlendioxid, daher könnte ein Methanausbruch („Clathratkanone“) eine schnelle globale Erwärmung auslösen oder diese noch schlimmer machen, wenn der Ausbruch selbst durch die globale Erwärmung verursacht würde.
Das wahrscheinlichste Zeichen eines solchen Methanausbruchs wäre ein plötzlicher Rückgang des Verhältnisses von Kohlenstoff-13 zu Kohlenstoff-12 in Sedimenten, da Methanclathrate einen niedrigen Kohlenstoff-13-Gehalt aufweisen; Allerdings müsste die Veränderung sehr groß sein, da auch andere Ereignisse den Anteil von Kohlenstoff-13 reduzieren können. [164]
Es wurde vermutet, dass Methanausbrüche mit „Clathratkanonen“ am Aussterben am Ende des Perms („dem Großen Sterben“) und am Paläozän-Eozän-Wärmemaximum beteiligt waren , das mit einem der kleineren Massenaussterben verbunden war.
Anoxische Ereignisse
Anoxische Ereignisse sind Situationen, in denen es in den mittleren und sogar oberen Schichten des Ozeans zu einem Mangel oder einem völligen Sauerstoffmangel kommt. Ihre Ursachen sind komplex und umstritten, aber alle bekannten Fälle werden mit einer schweren und anhaltenden globalen Erwärmung in Verbindung gebracht, die meist durch anhaltenden massiven Vulkanismus verursacht wird. [165]
Es wurde vermutet, dass anoxische Ereignisse das Ordovizium–Silurium , [166] [167] [168] spätes Devon , [169] [170] [171] Capitanium , [172] [173] [174] Permium verursachten oder dazu beitrugen. Trias- , [175] [176] [177] und Trias-Jura- Aussterben [178] sowie eine Reihe kleinerer Aussterben (wie Ireviken , Lundgreni , Mulde , Lau , Smithian-Spathium , Toarcian und Cenomanium-Turonium). Veranstaltungen). Andererseits gibt es ausgedehnte Schwarzschieferschichten aus der mittleren Kreidezeit, die auf anoxische Ereignisse hinweisen, aber nicht mit Massenaussterben in Verbindung gebracht werden.
Es wurde gezeigt, dass die Bioverfügbarkeit essentieller Spurenelemente (insbesondere Selen ) auf potenziell tödliche Tiefststände mit mindestens drei Massenaussterben in den Ozeanen zusammenfällt und wahrscheinlich dazu beigetragen hat, nämlich am Ende des Ordoviziums. während des mittleren und späten Devon und am Ende der Trias. In Zeiten niedriger Sauerstoffkonzentration wird sehr lösliches Selenat (Se 6+ ) in viel weniger lösliches Selenid (Se 2- ), elementares Se und Organo-Selen-Komplexe umgewandelt . Die Bioverfügbarkeit von Selen sank während dieser Aussterbeereignisse auf etwa 1 % der aktuellen ozeanischen Konzentration, ein Wert, der sich für viele noch lebende Organismen als tödlich erwiesen hat. [179]
Der britische Ozeanologe und Atmosphärenforscher Andrew Watson erklärte, dass das Holozän zwar viele Prozesse aufweist, die an jene erinnern, die zu früheren anoxischen Ereignissen beigetragen haben, dass es jedoch „Tausende von Jahren dauern würde, bis sich eine vollständige Anoxie im Ozean entwickelt“. [180]
Schwefelwasserstoff-Emissionen aus den Meeren
Kump, Pavlov und Arthur (2005) haben vorgeschlagen, dass während des Perm-Trias-Aussterbeereignisses die Erwärmung auch das ozeanische Gleichgewicht zwischen photosynthetisierendem Plankton und sulfatreduzierenden Tiefwasserbakterien störte , was zu massiven Schwefelwasserstoffemissionen führte , die das Leben auf beiden Landflächen vergifteten und das Meer und schwächte die Ozonschicht erheblich , wodurch ein Großteil des noch verbliebenen Lebens der tödlichen UV-Strahlung ausgesetzt wurde . [181] [182] [72]
Ozeanischer Umsturz
Ozeanischer Umsturz ist eine Störung der thermohalinen Zirkulation , die dazu führt, dass Oberflächenwasser (das aufgrund der Verdunstung salzhaltiger als Tiefenwasser ist) direkt nach unten sinkt, wodurch anoxisches Tiefenwasser an die Oberfläche gelangt und dadurch die meisten dort lebenden sauerstoffatmenden Organismen abgetötet werden Oberflächen- und Mitteltiefen. Es kann entweder zu Beginn oder am Ende einer Vereisung auftreten , obwohl ein Umkippen zu Beginn einer Vereisung gefährlicher ist, da in der vorangegangenen Warmzeit ein größeres Volumen an anoxischem Wasser entstanden ist. [183]
Im Gegensatz zu anderen ozeanischen Katastrophen wie Regressionen (Sinken des Meeresspiegels) und anoxischen Ereignissen hinterlassen Umstürze keine leicht erkennbaren „Signaturen“ im Gestein und sind theoretische Konsequenzen der Schlussfolgerungen der Forscher über andere Klima- und Meeresereignisse.
Es wurde vermutet, dass ozeanische Umwälzungen das Aussterben im späten Devon und im Perm-Trias verursachten oder dazu beitrugen .
Geomagnetische Umkehrung
Eine Theorie besagt, dass Perioden erhöhter geomagnetischer Umkehrungen das Erdmagnetfeld lange genug schwächen , um die Atmosphäre den Sonnenwinden auszusetzen, was dazu führt, dass Sauerstoffionen mit einer um drei bis vier Größenordnungen erhöhten Geschwindigkeit aus der Atmosphäre entweichen, was zu einem katastrophalen Rückgang des Sauerstoffs führt. [184]
Plattentektonik
Die Bewegung der Kontinente in bestimmte Konfigurationen kann auf verschiedene Weise zum Aussterben führen oder dazu beitragen: durch die Einleitung oder Beendigung von Eiszeiten ; durch veränderte Meeres- und Windströmungen und damit eine Veränderung des Klimas; durch die Öffnung von Seewegen oder Landbrücken, die zuvor isolierte Arten einer Konkurrenz aussetzen, an die sie schlecht angepasst sind (z. B. das Aussterben der meisten einheimischen Huftiere Südamerikas und aller großen Metatherianer nach der Schaffung einer Landbrücke zwischen Nord- und Südamerika). ). Gelegentlich entsteht durch Kontinentaldrift ein Superkontinent, der den größten Teil der Landfläche der Erde umfasst, was zusätzlich zu den oben aufgeführten Auswirkungen wahrscheinlich die Gesamtfläche des Festlandsockels (des artenreichsten Teils des Ozeans) verringert und a ausgedehntes, trockenes Kontinentalinneres, das extreme saisonale Schwankungen aufweisen kann.
Eine andere Theorie besagt, dass die Entstehung des Superkontinents Pangäa zum Massensterben am Ende des Perms beigetragen hat . Pangäa war am Übergang vom mittleren Perm zum späten Perm fast vollständig ausgebildet, und das Diagramm „Vielfalt der marinen Gattungen“ oben in diesem Artikel zeigt einen Grad des Aussterbens ab diesem Zeitpunkt, der möglicherweise für die Aufnahme in die „Marine-Gattungsvielfalt“ in Frage gekommen wäre. „Big Five“, wenn es nicht vom „Großen Sterben“ am Ende des Perm überschattet würde. [185]
Andere Hypothesen
.jpg?_x_tr_sl=en&_x_tr_tl=de&_x_tr_hl=de&_x_tr_pto=wapp)
Es wurden viele andere Hypothesen aufgestellt, beispielsweise die Ausbreitung einer neuen Krankheit oder die einfache Verdrängung der Konkurrenz nach einer besonders erfolgreichen biologischen Innovation. Aber alle wurden abgelehnt, meist aus einem der folgenden Gründe: Sie erfordern Ereignisse oder Prozesse, für die es keine Beweise gibt; sie gehen von Mechanismen aus, die den verfügbaren Beweisen widersprechen; Sie basieren auf anderen Theorien, die abgelehnt oder ersetzt wurden.

Wissenschaftler befürchten, dass menschliche Aktivitäten dazu führen könnten, dass mehr Pflanzen und Tiere aussterben als je zuvor. Neben vom Menschen verursachten Klimaveränderungen (siehe oben) könnten einige dieser Artensterben auch auf Überjagung, Überfischung, invasive Arten oder den Verlust von Lebensräumen zurückzuführen sein. In einer im Mai 2017 in Proceedings of the National Academy of Sciences veröffentlichten Studie wurde argumentiert, dass aufgrund anthropogener Ursachen wie Überbevölkerung und Überkonsum eine „biologische Vernichtung“ im Gange sei, die einem sechsten Massenaussterben ähnelt . Die Studie ergab, dass bis zu 50 % der einst auf der Erde lebenden Tierarten bereits ausgestorben waren, was auch die Existenzgrundlage des Menschen bedrohte. [187] [30]
Zukünftiges Aussterben/Sterilisieren der Biosphäre
Siehe auch:
Zukunft der Erde und
Die eventuelle Erwärmung und Ausdehnung der Sonne könnte zusammen mit dem eventuellen Rückgang des atmosphärischen Kohlendioxids tatsächlich zu einem noch größeren Massensterben führen, das möglicherweise sogar Mikroben auslöschen könnte (mit anderen Worten, die Erde wäre völlig steril): Aufstieg Die durch die expandierende Sonne verursachten globalen Temperaturen würden die Verwitterungsrate allmählich erhöhen, was wiederum der Atmosphäre immer mehr CO 2 entziehen würde. Wenn der CO2 – Gehalt zu niedrig wird (vielleicht auf 50 ppm), sterben die meisten Pflanzen aus, obwohl einfachere Pflanzen wie Gräser und Moose viel länger überleben können, bis der CO2- Gehalt auf 10 ppm sinkt. [188] [189]
Da alle photosynthetischen Organismen verschwunden sind, kann der Luftsauerstoff nicht mehr nachgefüllt werden und wird schließlich durch chemische Reaktionen in der Atmosphäre, möglicherweise durch Vulkanausbrüche, entfernt. Letztendlich führt der Sauerstoffverlust dazu, dass das gesamte verbleibende aerobe Leben durch Erstickung ausstirbt und nur einfache anaerobe Prokaryoten zurückbleiben . Wenn die Sonne in etwa einer Milliarde Jahren um 10 % heller wird, [188] wird die Erde einen feuchten Treibhauseffekt erleiden, der dazu führt, dass ihre Ozeane verdampfen, während der flüssige äußere Kern der Erde aufgrund der Ausdehnung des inneren Kerns abkühlt und das Erdmagnetfeld zum Absinken bringt abschalten. Ohne ein Magnetfeld werden geladene Teilchen von der Sonne die Atmosphäre erschöpfen und die Temperatur der Erde in 2,8 Milliarden Jahren weiter auf durchschnittlich etwa 420 K (147 °C, 296 °F) erhöhen, wodurch das letzte verbliebene Leben entsteht Die Erde wird aussterben. Dies ist der extremste Fall eines klimabedingten Aussterbens. Da dies erst spät im Leben der Sonne geschehen wird, wäre es das letzte Massenaussterben in der Erdgeschichte (wenn auch ein sehr langes Aussterben). [188] [189]
Auswirkungen und Erholung
Die Auswirkungen von Massenaussterben waren sehr unterschiedlich. Nach einem großen Aussterben überleben aufgrund ihrer Fähigkeit, in vielfältigen Lebensräumen zu leben, normalerweise nur Unkrautarten . [190] Später diversifizieren sich die Arten und besetzen leere Nischen. Im Allgemeinen dauert es Millionen von Jahren, bis sich die Artenvielfalt nach Aussterbeereignissen erholt. [191] Bei den schwersten Massenaussterben kann es 15 bis 30 Millionen Jahre dauern. [190]
Das schlimmste Ereignis im Phanerozoikum , das Aussterben im Perm und der Trias , zerstörte das Leben auf der Erde und tötete über 90 % der Arten. Das Leben schien sich nach dem Aussterben der PT schnell zu erholen, allerdings geschah dies meist in Form von Katastrophentaxa , wie dem robusten Lystrosaurus . Die neuesten Forschungsergebnisse zeigen, dass die spezialisierten Tiere, die komplexe Ökosysteme mit hoher Artenvielfalt, komplexen Nahrungsnetzen und einer Vielzahl von Nischen bildeten, viel länger brauchten, um sich zu erholen. Es wird angenommen, dass diese lange Erholung auf aufeinanderfolgende Aussterbewellen zurückzuführen war, die die Erholung behinderten, sowie auf anhaltenden Umweltstress, der bis in die frühe Trias anhielt. Neuere Forschungen deuten darauf hin, dass die Erholung erst zu Beginn der mittleren Trias, vier bis sechs Millionen Jahre nach dem Aussterben, einsetzte; [192] und einige Autoren schätzen, dass die Erholung erst 30 Millionen Jahre nach dem Aussterben der PT, also in der späten Trias, abgeschlossen war. [193] Nach dem Aussterben der PT kam es zu einer zunehmenden Provinzialisierung, wobei die Arten kleinere Verbreitungsgebiete besiedelten – was möglicherweise dazu führte, dass die etablierten Arten ihre Nischen verdrängten und die Voraussetzungen für eine eventuelle Neudiversifizierung geschaffen wurden. [194]
Die Auswirkungen des Massenaussterbens auf Pflanzen sind aufgrund der inhärenten Verzerrungen im Pflanzenfossilbestand etwas schwieriger zu quantifizieren. Einige Massenaussterben (wie das Ende des Perms) waren für Pflanzen gleichermaßen katastrophal, während andere, wie das Ende des Devons, die Flora nicht beeinträchtigten. [195]
Siehe auch
- Bioevent
- Elvis-Taxon
- Gefährdete Spezies
- Geologische Zeitskala
- Globales Katastrophenrisiko
- Holozänes Aussterben
- Aussterben der Menschheit
- Kačák-Ereignis
- Lazarus-Taxon
- Liste der Einschlagskrater auf der Erde
- Liste der größten Vulkanausbrüche
- Liste möglicher Einschlagstrukturen auf der Erde
- Medea-Hypothese
- Seltene Spezies
- Signor-Lipps-Effekt
- Schneeball-Erde
- Spekulative Entwicklung
- Das sechste Aussterben: Eine unnatürliche Geschichte (Sachbuch)
- Zeitleiste des Aussterbens im Holozän
- Quartäres Aussterben
Fußnoten
- ^ Die biologische Vielfalt nimmt schneller ab als jemals zuvor in der Geschichte der Menschheit. Die aktuellen Aussterberaten liegen beispielsweise etwa 100–1.000 Mal höher als die Basisrate, Tendenz steigend. [27]
- ^ „Das anhaltende sechste Massensterben stellt möglicherweise die schwerwiegendste Umweltbedrohung für den Fortbestand der Zivilisation dar, da es irreversibel ist. Tausende Populationen vom Aussterben bedrohter Wirbeltierarten sind in einem Jahrhundert verloren gegangen, was darauf hindeutet, dass das sechste Massensterben vom Menschen verursacht wurde.“ und beschleunigt sich. Die Beschleunigung der Aussterbekrise ist aufgrund des immer noch schnellen Wachstums der menschlichen Zahl und der Konsumraten sicher.“ — Ceballos, Ehrlich & Raven (2020) [32]
- ^ Gelöster Sauerstoff verbreitete sich weiter und drang in größere Tiefen vor; Die Entwicklung des Lebens an Land verringerte den Nährstoffabfluss und damit das Risiko von Eutrophierung und anoxischen Ereignissen . und die Meeresökosysteme wurden vielfältiger, so dass dieWahrscheinlichkeit einer Unterbrechung der Nahrungsketten geringer war.
- ^ Das früheste bekannte Basaltflutereignis ist dasjenige, das die Sibirischen Fallen hervorbrachte und mit dem Aussterben am Ende des Perms verbunden ist .
- ^ a bSpringe hoch zu: Einige der mit Überschwemmungsbasalten und Meeresspiegelabfällen verbundenen Aussterben waren deutlich geringer als die „großen“ Aussterben, aber immer noch viel größer als das Hintergrundaussterben.
No responses yet